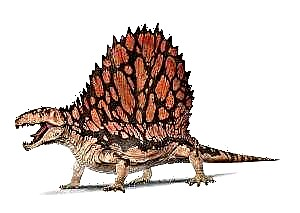
 Примерно 318 миллионов лет назад, во времена пермского периода, наша планета была населена звероподобными синапсидами. Эти древние существа отличались от современных рептилий не только размерами, но и строением зубного аппарата. Кроме резцов они имели еще и клыки, что может свидетельствовать об их хищнической натуре. Столь смелые заключения ученые смогли сделать на основании исследования многочисленных ископаемых останков, которые прекрасно сохранились в отложениях пермского периода. Впоследствии эти доисторические существа получили название диметродонтов.
Примерно 318 миллионов лет назад, во времена пермского периода, наша планета была населена звероподобными синапсидами. Эти древние существа отличались от современных рептилий не только размерами, но и строением зубного аппарата. Кроме резцов они имели еще и клыки, что может свидетельствовать об их хищнической натуре. Столь смелые заключения ученые смогли сделать на основании исследования многочисленных ископаемых останков, которые прекрасно сохранились в отложениях пермского периода. Впоследствии эти доисторические существа получили название диметродонтов.
Это были солидных размеров звероящеры, достигавшие в длину более 3,5 метров. Их отличительной особенностью являлся так называемый спинной парус. Это была высокая кожная складка, которая проходила вдоль всего позвоночника. Аналогичного рода выросты можно было наблюдать у некоторых видов древних амфибий и животных, к числу которых относятся динозавры и пеликозавры. По всей вероятности парус принимал активное участие в процессе терморегуляции тела этих животных. Если учесть, что температура окружающего воздуха, в те времена, была достаточно высокой, то холоднокровный диметродон легко мог погибнуть от перегрева, не будь у него паруса. Кроме того, столь оригинальное кожное образование могло исполнять роль вторичного полового признака, используемого диметродоном во время брачного сезона, а также применяться им для маскировки, при нахождении в густых тропических зарослях. Согласно иным гипотезам, кожную складку наш герой мог использовать в качестве паруса во время плавания.
Что касается образа жизни, то диметродоны жили в составе небольших групп. Взрослые особи отдавали предпочтение саваннам, а молодые предпочитали места, поросшие тропическим лесом. А вот в пустыне диметродон выжить не мог. Столь жаркий климат ему не подходил.
Это был свирепый и жестокий хищник, который нападал на всех представителей животного мира, с которыми мог справиться. Своими острыми, как бритва зубами и мощными челюстями, он легко рвал на куски тело своей жертвы.
По своему строению и образу жизни он больше походил на млекопитающих, нежели на рептилий. Ученые отнесли его к отряду пеликозавров, где он являлся самым многочисленным представителем.
По какой причине вымерли диметродоны, ученым доподлинно неизвестно. Возможно, она кроется в резких климатических изменениях, которых холоднокровные животные просто не смогли пережить. Согласно другой гипотезе, их вытеснили более совершенные существа.
Раннее изучение
Первоначально останки диметродона описал Эдвард Дринкер Коуп в 1870-ых. Он получил их вместе с коллекцией других пермских тетраподов, полученных из формации Красные грады в Техасе. Передали их Коупу коллекционер Якобс Болл, геолог У. Ф. Камминс и палеонтолог Чарльз Стернберг Большая часть этих образцов сейчас находится в Американском музее Естественной истории или в музее Уокера Чикакгского университета.
Стернберг отправил несколько своих образцов немецкому палеонтологу Фердинанду Бройли из Мюнхенского университета, однако он не так хорошо изучил их, как Коуп. Соперник Эдаврда, Чарльз Марш, также собрал несколько костей диметродона, однако он отдал их в музей Уокера.
Впервые наименование Dimetrodon использовал Коуп в 1878 году, выделив три вида — D. incisivus, D. rectiformis и D. gigas.
Одна из самых известных картин с диметродоном
Однако первое описание останков диметродона было в 1875 году, когда Коуп описал клепсидропса C. limbatus. Это животное было обнаружено в той же местности, что и диметродон, и в конце XIX-начале ХХ века большинство останков пеликозавров приписывали диметродону или клепсидопсу. В 1940 году вышла публикация, в которой говорилось, что C. limbatus является на самом деле видом диметродона.
Первым описанным синапсидом с парусом был клепсидропс C. natalis, также выделенный Коупом. Он посчитал парус плавником и сравнивал с гребнем ящерицы василиска. Паруса D. incisivus и D. gigas не сохранились, однако у образца D. rectiformi сохранились удлинённые невральные шипы. Однако Коуп прокомментировал в 1886 году, что назначение паруса сложно представить. По его словам, если животное не вело водный образ жизни, то парус или плавник мешал бы движениям, а конечности и когти не были достаточно большими, чтобы вести древесный образ жизни, как у василиска.
ХХ век
В начале ХХ века Э. Кейс провёл серьёзное исследование диметродона, выделив несколько новых видов. Ему помог с финансированием Институт Карнеги, предоставив также материал из различных музеев Америки. Многие образцы были описаны Копом, прославившимся тем, что описывал целые рода лишь по фрагментам, однако он не обратил на эти останки много внимания.
Начиная с концов 1920-ых Альфред Ромер провёл повторное изучение многих образцов демитродона, выделив ещё несколько видов. В 1940 году Ромер и Ллевеллин Прайс провели "Обзор пеликозавров", в котором просмотрели большинство синапсид, описанных Копом. Большинство результатов этого исследования актуальны до сих пор.
Старая реконструкция вида Dimetrodon incisivus
После публикации Ромера и Прайса последовало открытие многочисленных образцов диметродона вне Оклахомы и Техаса. Так, в 1966 году были обнаружены небольшие фрагменты на территории Юты, а в 1969 году останки нашли в Аризоне. В 1975 году Олсон сообщил о находке диметродона в Огайо. В 1977 Берман описал на основе материала из Нью-Мексико вид D. occidentalis ("западный"), к которому также отнесли останки из Юты и Аризоны.
До этих открытий считалось, что Техас и Оклахому от остальных территорий отделял Серединноконтинентальный морской путь, благодаря чему в западной Северной Америке обитал меньший сфенакодон. Новые находки хоть и не опровергают наличие внутреннего моря, но указывают на его ограниченность и то, что оно не было преградой для расселения диметродона.
Описание
У диметродона была довольно большая голова с крепкими челюстями, усеянными острыми зубами. Вероятно, он был активным охотником: охотился на амфибий, пресмыкающихся и рыбу. Своими передними зубами диметродон держал жертву и разрывал ее на части. Задние зубы были загнуты назад, с их помощью животное разрывало на части мелких существ и пережевывало большие куски мяса. Туловище диметродона имело бочкообразную форму. Наиболее яркой чертой диметродона является парус из натянутой на выросты спинных позвонков кожи. Подобные паруса развились у различных доисторических животных (амфибии платигистрикса, пеликозавров эдафозавра и секодонтозавра, динозавра спинозавра) и служили терморегуляторами. По другим версиям парус использовался в брачных играх, служил камуфляжем среди вертикальных стеблей растений или настоящим парусом при плавании. «Парус» развивался постепенно в течение жизни, известны находки молодых особей диметродона с невысокими остистыми отростками. Молодые особи, по-видимому, обитали в зарослях по берегам водоемов, как и взрослые. Тем не менее, бытующее в популярных книгах изображение диметродона на фоне пустыни представляется нереальным — не являясь рептилией он не мог бы выжить в жаркой сухой местности.
Череп
Череп диметродона высокий, сжатый с боков. Премаксилла отделена от челюсти глубокой диастемой. Передний край сильно выпуклый. Небольшие ноздри находятся в передней части морды. Слёзная кость не доходит до ноздрей. Глазницы находятся в задней части черепа. На премаксилле находится три зуба, второй зуб верхней челюсти имеет клыковидную форму, увеличен, изогнут, с зазубренными краями. Зубцы очень тонкие, как у альбертозавра. У их оснований находились небольшие пустоты, снижающие нагрузку на зубы, но не спасающая от повреждений зубцов. У D. teutonis не было зазубрин, однако края были такие же острые.. Нижний клык входит в диастему между премаксиллой и челюстью. Зубы обеих челюстей перед клыками редуцированны, за ними - постепенно уменьшаются в размерах. По форме зубы диметродона и его родственников напоминают капли, что помогает отличить сфенакодотво от остальных ранних синапсид.
Исследования 2014 года показали, что диметродоны проводили своего рода гонку вооружений. Мелкие D. milleri не имели зазубрин на зубах, так как охотились на мелкую добычу. По мере увеличения видового разнообразия и размеров, зубцы изменяли свою форму. У D. limbatus зубцы имели режущую форму, как у секодонтозавра. У крупного D. grandis зубцы имели такую же форму, как у акул и тероподов. Таким образом, диметродоны не только увеличивали размеры по мере эволюции, но и изменяли привычные приспособления для охоты.
От завропсид диметродон отличается наличием инфратемпоральной фенестры. У рептилий было две фенестры, либо они отсутствовали вообще, в то время как у синапсид было всего одно такое отверстие. У диметродона были необычные признаки перехода от ранних тетропод к млекопитающим, как гребни на задней части нижней челюсти и внутри носовой полости.
На внутренней части носовой полости находились специальные гребни, насотурбины. Они могли поддерживать хрящи, увеличивающие площадь обонятельного эпителия. Эти гребни меньше, чем у млекопитающих и более поздних синапсид, у которых насотурбины являются возможным признаком теплокровности. На них могла находиться слизистая оболочка, нагревающая и увлажняющая поступающий воздух. Таким образом, диметродон был частично теплокровным животным.
Ещё одной особенностью диметродона является выступ в задней части челюсти, называемый отражённой пластинкой. Она находится на суставной кости, соединённой с квадратной костью, вместе образующими челюстной сустав. У более поздних синапсид отростки суставной и квадратной кости отсоединились от челюстного сустава, образовав кость среднего уха - молоточек. отражённая пластинка позднее развилась в барабанное кольцо, поддерживающее барабанную перепонку у современных млекопитающих.
Хвост
Долгое время диметродона представляли как животное с коротким хвостом, так как всего было известно 11 ближайших к телу хвостовых позвонков, которые сильно сужались по мере отдаления от таза, а у первых описанных останков хвост вообще отсутствовал, либо был не в лучшем состоянии. Только в 1927 году был открыт полный хвост диметродона, состоящий из 50 позвонков. Он составлял большую часть длины тела и служил балансиром при передвижении.
Парус
Скелет D. loomisi
Одна из самых ярких и выдающихся черт диметродона - высокие остистые отростки спинных и шейных позвонков. Со времен открытия рода их изображали по-разному: как просто торчащие шипы, так и полностью обтянутый кожей «парус», или даже горб. Парус достигал в высоту до одного метра. Верхушки невральных шипов были покрыты роговыми чехликами. Каждый невральный шип имеет уникальную форму, такая дифференциация даже получила название "диметродонтовая". Возле тела позвонка шипы имеют прямоугольную форму, по мере удаления от него принимая форму восьмёрки. Считается, что такая форма укрепляла отростки и мешала переломам. Известна одна особь D. giganhomogenes с полностью прямоугольными невральными шипами, однако возле центра всё ещё присутствуют следы "восьмёрок". Скорее всего, эти изменения связаны с возрастом особи. Микроанатомия каждого шипа позволяет разглядеть место прикрепления к мышцам и место перехода в парус. Нижняя, проксимальная часть шипа имеет шероховатую поверхность. Возможно, к ней крепились эпаксальные и гипаксальные мышцы, а также сеть соединительной ткани, так называемые волокна шарпи. Дистальная часть шипов гладкая, однако надкостница пронизана многочисленными бороздками, возможно, при жизни в них располагались кровеносные сосуды. Многослойная пластинчатая кость, составляющая большую часть поперечного сечения невральных шипов, содержит много линий роста, по которым можно определить возраст каждой особи на момент смерти.
Через все позвонки проходит необычный желобок. Раньше считалось, что в нём располагались кровеносные сосуды, но так как их следы отсутствуют внутри костей, возможно, что желобок предназначался для чего-то другого, а количество сосудов в парусе было гораздо меньше, чем считалось.
Современная реконструкция скелета диметродона. Автор - Скотт Хартман
Изучение патологий показало, что некоторые невральныеные шипы сломались, а затем исцелились. Это наводит на мысль, что по крайней мере часть отростков была обтянута парусом (или подобной тканью), которая удерживала их на месте после повреждения, в следствии чего они могли заживать. Но они также заметили, что вершины шипов часто согнуты, иногда очень сильно, что предполагает, что вершина шипов не входила в парус. Дальнейшее подтверждение этой конфигурации «выход из паруса» происходит из поверхностной текстуры отростков. Они были шероховатыми в том месте, где крепились к мышцам спины, далее становясь все более гладкими, как раз там, где они, скорее всего, становились шипами, торчащими из паруса. Выросшая на месте переломов кортикальная кость сильно васкуляризирована, что говорит о наличие определённого числа мягких тканей и сосудов на парусе.
Отпечатки кожи диметродона неизвестны, поэтому говорить о какой-то определённой текстуре сложно. У более поздних синапсидов, вроде эстемменозуха, кожа была гладкой, с большим количеством желёз. Однако у более примитивных ранних варанопсеид, таких как аскендонан, удалось обнаружить отпечатки чешуи. Не исключено, что диметродон имел чешуйчатую текстуру на нижней стороне тела, а с боков и сверху его кожа была гладкой, как у терапсид.
Походка
Диметродон традиционно изображается с развалистой "ящеричной" походкой с волочащимся по земле брюхом, однако недавно были обнаружены дорожки, принадлежащие диметродону или близкому к нему синапсиду, показывающие животное, передвигающееся с более выпрямленными ногами, держа живот и хвост полностью свободными от земли.
Конечно, диметродон мог разлечься, когда захочет. Однако во время ходьбы и бега его конечности всё-таки имели полувыпрямленное положение, из-за чего диметродон мог быть быстрее своих жертв (амфибий и более мелких синапсид).
Хорошо известные виды
- D. teutonis Reisz & Berman, 2001. Нижние слои верхнего красного лежня (вольфкамп), Германия, Бромакер и Россия. Самый маленький диметродон, весил около 24 кг. Единственный достоверно известный вид за пределами Северной Америки. Обладал высоким парусом. Был привязан к сухопутной биоте.
Скелет Dimetrodon milleri
- D. milleri Romer 1937. Сакмарский ярус, длина до 174 см, формация Putnam, Техас. Известен по двум скелета: почти полному MCZ 1365 и более крупному, но не так хорошо сохранившемуся MCZ 1367. Самый ранний вид диметродона из Техаса. Отличается от остальных видов строением невральных шипов: у D. milleri они имеют округлую форму, в то время как у остальных видов они напоминают восьмёрку в поперечнике. У этого вида позвонки также короче, чем у остальных. Череп высокий, морда короткая. Похожее строение имеют также D. booneorum, D. limbatus и D. grandis, возможно, что D. milleri был их предком. Близок к D. occidentalis. Син.:Clepsydrops natalis Cope, 1887.
- D. natalis Cope 1877. Сакмарский ярус, самый мелкий американский вид. Изображается с низким трапецевидным парусом, однако настоящая форма неизвестна. Длина черепа около 14 см, вес до 37 кг. Техас. Череп низкий, на верхней челюсти находились клыкообразные зазубренные зубы. Обнаружен недалеко от тела крупного D. limbatus.
Скелет D. incisivus
- D. limbatus Cope 1877. Сакмарский и Артинский ярус - длина черепа до 40 см, общая длина до 2,6 м, из формации Admiral и Bell Plains в Техасе. Самый первый известный синапсид с парусом. Часто изображается в литературе. Первоначально был описан как Clepsydrops limbatus, к диметродону его отнесли Ромер и Прайс в 1940 году. Син.:Clepsydrops limbatus Cope, 1877, ? Dimetrodon incisivus Cope, 1878, Dimetrodon rectiformis Cope, 1878, Dimetrodon semiradicatus Cope, 1881.
- D. incisivus Cope, 1878 — один из первых видов, иногда считается типовым. Возможный синоним D. limbatus.
- D. booneorum Romer 1937 — Артинский ярус — длина до 2,2 метра, Техас. Описан Ромером в 1937 году.
- D. gigashomogenes Case 1907. Кунгурский ярус. Достигал в высоту до 3,3 метров. Череп короткий и относительно высокий. Один из предков D. angelensis. Обнаружен в формации Арройо. Назван Кейсом ещё в 1907 году, до сих пор валиден.
Скелет D. grandis Case, 1907
- D. grandis Case, 1907. Ранний Кунгурский ярус. Достигал в длину до 3,2 м. Череп низкий, длиной 50 см. Имел всего четыре предклыковых зуба. Обнаружен в формации формации Арройо, Техас. Син.: Theropleura grandis Case, 1907, Bathyglyptus theodori Case, 1911, ? Dimetrodon gigas Cope, 1878, Dimetrodon maximus Romer, 1936, ? Dimetrodon cf. gigas grandis Sternberg, 1942.
- D. loomisi Romer 1937 . Кунгурский ярус. Вырастал до 2,5 метров. Обнаружен в формации Арройо, Техас. Отличается довольно низким черепом и зазубренной формой паруса.
- D. angelensis Olson 1962. Ранняя уфимская эпоха (поздняя Кунгурская). Последний и самый крупный известный вид. При жизни вырастал до 4—4,5 метров. Обнаружен в формации Сан-Анжело в Техасе. Череп длинный, до 50 см, и низкий, верхние клыки длинные тонкие. Все образцы плохо сохранились. Син.: ? Eosyodon hudsoni Olson, 1962 (nomen dubium), ? Steppesaurus gurleyi Olson & Beerbower, 1953.
- D. borealis Leidy, 1854. 270 млн лет назад, Остров Принца Эдуарда. Возможный вид, известный также как батигнат. Возраст останков в этом районе подтвердился после исследования остатков растений. Позднее была найдена вся голова батигната. Длина черепа всего 40-45 см.
Отпечаток ноги диметродона
- D. occidentalis Berman 1977 — единственный диметродон из формации Або/Катлер Нью-Мексико. Прелполагается, что достигал в длину 1,5 метра. Название означает "Западный диметродон". Известен по одному небольшому скелету. Предположительно связан с D. milleri.
- D. gigas Cope, 1878. Артинский и Кунгурский ярусы перми. Первоначально был описан как Clepsydrops gigas, однако позже его реклассифицировали как диметродона. К виду относится несколько хорошо сохранившихся образцов. Также рассматривается как синоним D. grandis.
- D. macrospondylus Case, 1907 — описан Копом как Clepsydrops macrospondylus, к диметродону определён Кейсом.
ЧЕМ ПИТАЛСЯ
Несмотря на то, что кости черепа диметродона были достаточно тонкими, своими крепкими челюстями, усеянными острыми зубами, он намертво впивался в жертву. Зубы были разной величины, коренные зубы были загнуты назад. Своими длинными передними зубами он кусал жертву, как это делают современные львы. Диметродон крепко хватал добычу. Учёные считают, что этот хищник охотился на амфибий, рептилий и рыбу. Своми передними зубами диметродон держал жертву и разрывал её на части. Задние зубы были загнуты назад, с их помощью животное разрывало на части мелких сумчатых и пережёвывало большие куски мяса.
ОБРАЗ ЖИЗНИ
Диметродон был одним из крупнейших и самых свирепых хищников раннего Пермского периода. Эти животные исчезли с лица земли ещё до появления первых динозавров.
Палеонтологи, исследовавшие останки диметродонов, пришли к выводу, что это были хорошо вооружённые и свирепые хищники. Диметродон был величиной с современный автомобиль. У него были короткие мощные конечности, поэтому учёные полагают, что диметродон передвигался по земле подобно современным ящерицам.
Вероятно, он был довольно медлительным животным. Нет точных данных о том, сколько весил диметродон, однако предполагают, что его масса была довольно значительной. Большой парусообразный нарост на спине придавал ему устрашающий вид. Считается, что с помощью этого „паруса" животное могло регулировать температуру своего тела.
Утром диметродон грелся на солнце, через парус тепло передавалось другим частям тела этого ящера, поэтому он согревался скорее вялых потенциальных жертв. Чтобы охладиться, диметродону было достаточно погрузить свой парус в воду. Учёным не известно, почему исчезли диметродоны.
ОБЩИЕ ПОЛОЖЕНИЯ. ОПИСАНИЕ
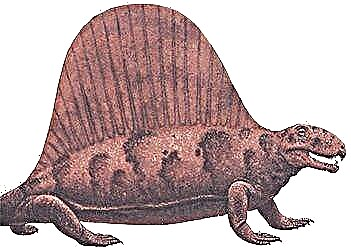
Пермь, 280 млн лет назад
Северная Америка
Длина 3,5 м
Это самый известный из пеликозавров, древнейших звероподобных ящеров. Тело у него было крепкое, ноги короткие, челюсти сильные, с острыми зубами. На спине - высокий кожистый парус, который поддерживали остистые отростки позвонков. Его функции точно не известны. Большинство учёных считает, что «парус» помогал поддерживать оптимальную температуру тела: на солнце кровь в кровеносных сосудах паруса нагревалась, а в тени - охлаждалась. Хотя, возможно, это был ярко окрашенный гребень для привлечения половых партнёров.
ИНТЕРЕСНАЯ ИНФОРМАЦИЯ. ИЗВЕСТНО ЛИ ВАМ, ЧТО.
- Название диметродон имеет греческое происхождение. Она состоит из двух слов „dimitro" - „то, что существует в двух измерениях", и „don", то есть „зуб".
- Палеонтологи проводили очень длительные исследования останков „паруса" диметродона, на основании которых пришли к выводу, что парус служил для регулирования температуры тела этого ископаемого животного.
- Кожа „паруса", которая была на спине диметродона, вероятно, была водонепроницаемая, как у современных пресмыкающихся.
ХАРАКТЕРНЫЕ ОСОБЕННОСТИ ДИМЕТРОДОНА
Парус: парусообразный нарост располагался на спине диметродона от шеи и до таза Учёные считают, что парус был своеобразной системой температурного контроля. Утром диметродон грелся в лучах солнца, солнечные лучи согревали парус, а через него тепло передавалось в другие органы тела животного. Вероятно, чтобы избежать перегрева, диметродон погружал парус в воду. Согласно другой версии, парус могл иметь и другую функцию, например, служил половому признаку - у самцов могло быть больше и ярче парус, чем у самок.
Зубы: длинные, крепкие глазные зубы использовались для захвата добычи и её разрывания на части. Короткие коренные зубы были загнуты назад, с их помощью диметродон прочно удерживал добычу и пережёвывал куски мяса.
Череп: глава была довольно большими. Отверстие, расположенное позади глазниц, уменьшало массу черепа. К задней части черепа крепились сильные мышцы.
Ноги: и задние, и передние конечности этого животного были короткими и массивными. Они должны были выдерживать вес тела этого гигантского ящера. Кроме того, крепкие мышцы задних конечностей удерживали длинный хвост.

- Находки окаменелостей археоптерикса
ГДЕ И КОГДА ЖИЛ
В настоящее время найдено 6 окаменелостей археоптерикса. Все - в Баварии. Во времена, когда жили археоптериксы, территория Германии была частью континента, который выглядел совершенно иначе, и находился в тропиках. На основе определения геологического возраста сланцев, в которых были найдены окаменелости, стало известно, что археоптерикс жил в Верхнеюрском периоде, то есть около 150 миллионов лет назад.
Место обитания
Большая часть ископаемых останков найдена на территории США, но также их можно встретить и в Германии (в начале пермского периода данные территории были объединены в континент). Диметродоны селились неподалеку от водоемов, но места их обитания менялись по мере взросления особей: детеныши предпочитали болотистую местность с густой растительностью, юное поколение выбирало берега озер, а матерые особи — широкие речные долины. Возможно, диметродоны вели полуводный образ жизни и хорошо плавали.
Особенности внешнего вида
Название «диметродон» означает «зубы двух типов». В пасти животного помимо мелких зубов располагались клыки и резцы (такая их дифференциация присуща млекопитающим). От рептилий диметродону достались особенности строения конечностей, которые были широко расставлены по бокам, а не вертикально под туловищем, а также холоднокровие. Температура его тела зависела от окружающей среды. Помимо паруса, характерной чертой животного был очень длинный хвост, состоявший из не менее чем 50 позвонков. Размеры диметродонов в зависимости от их вида могли сильно варьироваться — разброс по длине тела составлял от 0,6 до 4,6 м.

Череп синапсид известен височными впадинами. Они располагались по одной с каждой стороны, позади и чуть ниже глазниц. Впадины служили для крепления челюстных мышц. Их наличие делало укус синапсид более эффективным по сравнению с возможностями земноводных, у которых такая особенность строения черепа отсутствовала.
Особенности строения
Диметродон имел спинной парус, состоявший из длинных костных отростков позвонков, обтянутых кожей. Он мог выполнять терморегуляционную функцию, быстро нагреваясь на солнце. Ученые подсчитали, что без паруса температура тела взрослого диметродона поднялась бы на 6° за 3 ч 40 мин, а с ним — за 1 ч 20 мин. В тени кожаное украшение быстро отдавало тепло, спасая животное от перегрева. Кроме того, парус мог использоваться во время брачных игр для привлечения самок (предполагается, что у самцов такой спинной гребень был более развит). Он формировался постепенно, по мере взросления диметродона.

Доминирующий хищник
Диметродон считается самым крупным наземным хищником своего периода. Он мог охотиться на любых животных, обитавших с ним по соседству. Предполагается, что диметродон обладал развитым обонянием. У этих животных был выражен половой диморфизм, то есть внешне самки и самцы имели различия, не связанные с первичными половыми признаками (например, женские особи могли быть меньших размеров). Точно не известно, как жили диметродоны: группами или поодиночке. Не исключено, что самцы могли проявлять агрессию по отношению друг к другу.











