| Название вида: | Белый аист |
| Латинское название: | Ciconia ciconia (Linnaeus, 1758) |
| Английское название: | White stork |
| Французское название: | Cigogne blanche |
| Немецкое название: | WeifJstorch |
| Латинские синонимы: | для подвида ciconia: Ciconia alba Bechstein, 1793, Ciconia albescens C.L. Brehm, 1831, Ciconia nivea C.L. Brehm, 1831, Ciconia Candida C.L. Brehm, 1831, Ciconia major C.L. Brehm, 1855, для подвида asiatica: Ciconia asiatica Severtzov, 1873, Ciconia orientalis Severtzov, 1875 |
| Отряд: | Аистообразные (Ciconiformes) |
| Семейство: | Аистовые (Ciconiidae) |
| Род: | Аисты (Ciconia Brisson, 1760) |
| Статус: | Гнездящийся мигрирующий вид. |
Общая характеристика и полевые признаки
Крупная птица с длинными ногами, шеей и клювом. Длина тела 100-115 см, размах крыльев 155-165 см, вес взрослой птицы от 2,5 до 4,5 кг. Самцы несколько крупнее самок, но внешне они практически неотличимы. Оперение белое, маховые чёрные. Клюв и ноги красные. При наблюдении за летящей птицей, обращают на себя внимание вытянутые шея и ноги, контрастное чёрно-белое оперение. Ходит по земле, немного покачивая головой в такт движению. На гнёздах или присадах может подолгу стоять на одной ноге, втянув шею в оперение туловища. Часто использует парящий полёт, практически без взмахов крыльев способен подниматься в восходящих потоках воздуха. При резком снижении и посадке — немного прижимает крылья к туловищу и выставляет ноги вперёд. Стаи образуются во время перелёта, их формируют также негнездящиеся птицы в ходе кочёвок в поздневесенний и летний периоды. Строго порядка в летящих стаях нет. При снижении из восходящего потока птицы одна за другой скользят вниз. От чёрного аиста отличается белым оперением, от журавлей и цапель — красным цветом клюва и ног. В отличие от цапель, в полете вытягивает, а не складывает шею.
Голос. Основа звукового общения белых аистов — трещание клювом. Изредка можно услышать шипение. Звуковой репертуар птенцов более разнообразен. Крик выпрашивающего корм аистёнка напоминает протяжное мяуканье. Первая часть этого крика имеет более высокий тон, вторая — низкий. У птенцов на гнезде можно услышать также громкий писк и шипение, уже в первые недели жизни птенцы пытаются трещать клювом.
Описание
Окраска. Взрослые самец и самка. Сезонных различий в окраске нет. Большая часть оперения белая, первостепенные маховые, наружные второстепенные, плечевые и часть кроющих перьев предплечья чёрные с металлическим блеском. Наружные опахала второстепенных маховых имеют сероватые поля вдоль ствола (признак варьирует, виден обычно только с близкого расстояния). Перья на шее и груди несколько удлинены, возбуждённые птицы (например, во время токования) часто их распушивают. Клюв и ноги ярко-красные. Голая кожа вокруг глаза и передняя часть кожи подбородка чёрные. Радужина глаза бурая.
Первый пуховой наряд. Птенец после вылупления покрыт редким и коротким серовато-белым пухом. Ноги розоватые, через несколько дней становятся серовато-чёрными. Клюв и кожа вокруг глаз чёрные, на подбородке кожа красноватая, радужина тёмная. Второй пуховой наряд. Пух чисто белый, более густой и длинный. Сменяет первый примерно через неделю.
Гнездовой наряд. Молодая птица по окраске похожа на взрослую, но чёрный цвет в оперении заменён буроватым, без блеска. Клюв и ноги тёмно-бурые, ко времени вылета птенцов из гнёзд они обычно становятся красновато-бурыми, но нередко можно увидеть и летающих слетков с чёрным клювом или бурым с черноватой вершиной. Радужина глаза серая.
Строение и размеры
Как правило, публикуют промеры различных частей тела аистов, не разделяя выборку на половые группы. Длина крыла номинативного подвида белого аиста при таком подходе для территории бывш. СССР составляет, по 6 особям, 585— 605 мм (Спангенберг, 1951), для Украины (Смогоржевський, 1979) — 534-574 мм. Последний автор сообщает также, что длина хвоста колеблется в пределах 206-232 мм, клюва −156-195, а цевки — 193-227 мм. Ревизия коллекций Зоомузея Киевского национального университета и Национального научно-природоведческого музея Украины дал следующие результаты: длина крыла (п = 14) — 513-587 мм, при среднем значении 559,9±5,8 мм, хвоста (п = 11) — 201-232, в среднем 222,5±4,2, клюва (п = 12) — 150-192, в среднем 166,4±3,5, цевки (п = 14) — 187-217, в среднем 201,4±2,5 мм (ориг.). Для азиатского белого аиста длина крыла по 9 измеренным особям оказалась 550-640, в среднем 589 мм.
Размеры белого аиста по половым группам и подвидам для различных территорий приводятся в табл. 31.
| Параметр | Самцы | Самки | Источник | ||||
|---|---|---|---|---|---|---|---|
| n | lim | M | n | lim | M | ||
| Ciconia ciconia ciconia. Европа | |||||||
| Длина крыла | — | 530–630 | — | — | 530–590 | — | Witherby et al., 1939 |
| Длина хвоста | — | 215–240 | — | — | 215–240 | — | Witherby et al., 1939 |
| Длина клюва | — | 150–190 | — | — | 140–170 | — | Witherby et al., 1939 |
| Длина цевки | — | 195–240 | — | — | 195–240 | — | Witherby et al., 1939 |
| Длина крыла | 18 | 556–598 | 576 | 15 | 543–582 | 558 | Hancock et al., 1992 |
| Длина хвоста | 18 | 221–268 | 247 | 15 | 218–256 | 237 | Hancock et al., 1992 |
| Длина клюва | 18 | 157–198 | 179 | 15 | 155–180 | 164 | Hancock et al., 1992 |
| Длина цевки | 18 | 191–230 | 214 | 15 | 184–211 | 197 | Hancock et al., 1992 |
| Ciconia ciconia asiatica. Средняя Азия | |||||||
| Длина крыла | 18 | 581–615 | 596 | 9 | 548–596 | 577 | Hancock et al., 1992 |
| Длина клюва | 18 | 188–223 | 204 | 9 | 178–196 | 187 | Hancock et al., 1992 |
| Длина цевки | 18 | 213–247 | 234 | 9 | 211–234 | 220 | Hancock et al., 1992 |
Формула крыла (не считая рудиментарного первого махового) IV?III?V-I-VI. Наружные опахала II и IV первостепенных маховых имеют вырезки. Хвост слегка закруглён, рулевых перьев 12. Клюв длинный, прямой, суживающийся к вершине. Ноздри длинные, щелевидные. Вес 41 самца из Вост. Пруссии 2 900-4 400 г (в среднем 3 571), 27 самок — 2 700-3 900 г (3 325). Вес на протяжении лета немного увеличивается. Средний вес 14 самцов в июне 3 341 г, 14 самок — 3 150 г, в июле-августе 12 самцов весили в среднем 3 970 г, 12 самок — 3 521 г (Steinbacher, 1936).
Самец, таким образом, несколько крупнее самки, имеет более длинный и массивный клюв. Кроме того, клюв у самца имеет несколько иную форму: подклювье перед вершиной слегка изогнуто вверх, у самки же клюв прямой (Bauer, Glutz von Blotzheim, 1966, Creutz, 1988). По длине клюва можно определить пол 67% птиц с вероятностью ошибки не более 5% (Post et al., 1991). Возможно также индивидуальное узнавание птиц по рисунку чёрного пятна на подбородке (Fangrath, Helb, 2005).
Линька
Изучена недостаточно. У молодых птиц полная постювенильная линька начинается, в зависимости от обстоятельств, с декабря по май первого года жизни. У взрослых птиц полная линька занимает большую часть года. Первостепенные маховые сменяются в нерегулярной последовательности на протяжении гнездового периода, некоторые зимой (Stresemann, Stresemann, 1966).
Более детально линька маховых перьев прослежена на 5 аистах, содержавшихся в питомнике в Швейцарии (Bloesch et al., 1977). Рост пера происходит с линейной скоростью. Первостепенные маховые растут по 8-9 мм в день, второстепенные — 6,5-6,9 мм. На смену махового пера уходит от 50-55 до 65-75 дней. У находившихся под наблюдением птиц в год сменялось по 6 первостепенных маховых и 13 второстепенных на обоих крыльях. Продолжительность ношения различных перьев разная, для первостепенных маховых она колебалась от 1,2 до 2,5 лет. Смена перьев идёт ступенчато. У первостепенных маховых она начинается с XI, у второстепенных — с нескольких точек. Циклы линьки начинаются со второго года жизни, их окончательный ход устанавливается лишь к 4-5 году. Во время первой-третьей линьки смена перьев начиналась в марте-апреле, в дальнейшем — в середине мая и продолжалась до начала ноября. Большая часть перьев сменялась в летние месяцы между началом насиживания и отлётом.
Совмещение линьки и гнездования может быть связано с тем, что у белого аиста в это время нагрузка на крылья гораздо меньше, чем во время продолжительной миграции или при кочевой жизни на зимовках (Creutz, 1988).
Подвидовая систематика
Выделяют 2 подвида, отличающихся размерами и формой клюва:
1.Cicortia cicottia ciconia
Ardea ciconia Linnaeus, 1758, Syst. Nat., ed. 10, c. 142, Швеция.
Более мелкая форма. Длина крыла самцов 545-600 мм, длина цевки 188-226 мм, длина клюва 150-200 мм. Клюв менее резко скошен к вершине (Степанян, 2003). Распространён в Европе, Сев. Африке, Зап. Азии.
2.Ciconia ciconia asiatica
Ciconia alba asiatica Severtzov, 1873, Изв. Имп. о-ва любителей естествознания, антропологии и этнографии, 8, вып. 2, с. 145, Туркестан.
Более крупная форма. Длина крыла самцов 580-630 мм, длина цевки 200-240 мм, длина клюва 184-235 мм. Клюв, в особенности подклювье, более резко скошен к вершине (Степанян, 2003). Обитает на территории Узбекистана, Казахстана, Таджикистана и Киргизии.
Распространение
Гнездовой ареал. Европа, Сев.-Зап. Африка, Зап. и Средняя Азия .

Рисунок 78. Область распространения белого аиста:
а — область гнездования, б — районы зимовок, в — основные направления осеннего пролёта, г — направления экспансии.
Европейский подвид распространён на большей части Европы от Пиренейского п-ова до Поволжья и Закавказья. На север его ареал доходит до Дании, Юж. Швеции, Эстонии, северо-запада России. Во Франции аисты обитают лишь в нескольких провинциях, поэтому места гнездования в Испании, Португалии, Зап. Франции и Сев.-Зап. Африке оказываются оторванными от основного европейского ареала. Однако, в связи с продолжающимся расселением, вполне вероятно смыкание этих двух частей ареала. В Сев.-Зап. Африке белый аист гнездится в Марокко, на севере Алжира и Туниса. В Зап. Азии — в Турции, Сирии, Ливане, Израиле, Ираке, Иране, в Закавказье — на юге Грузии, в Армении, Азербайджане, а также в Республике Дагестан РФ. Известны случаи размножения также на местах зимовки в Юж. Африке (Broekhuysen, 1965, 1971, Broekhuysen, Uys, 1966, Hancock et al., 1992). В 2004 г. отмечена попытка гнездования на северо-востоке Англии в графстве Йоркшир. Это первый случай размножения белого аиста в Великобритании после 1416 г., когда птицы гнездились на кафедральном соборе в Эдинбурге.
В России белый аист издавна населяет территорию Калиниградской обл. В других областях он появился сравнительно недавно, расширяя гнездовой ареал в восточном и северо-восточном направлениях. Первые случаи гнездования в современных границах Ленинградской и Московской обл. отмечались ещё в конце XIX в. (Мальчевский, Пукинский, 1983, Зубакин и др., 1992). К началу XX в. белый аист начал гнездиться в Псковской, Тверской и Калужской обл. (Зарудный, 1910, Филатов, 1915, Бианки, 1922). К этому времени он уже был достаточно обычным в западных районах Смоленской (Граве, 1912, 1926) и южной части Брянской обл. (Федосов, 1959). Расселение на новые территории имело волнообразный характер. Особенно интенсивным освоение новых местностей отмечено в 1970-1990-х гг. В настоящее время на территории России северную и восточную границу регулярного гнездования восточноевропейской популяции можно условно провести по линии Санкт-Петербург — Волхов — Тихвин — Ярославль — Липецк — Воронеж — граница Ростовской обл. и Украины .

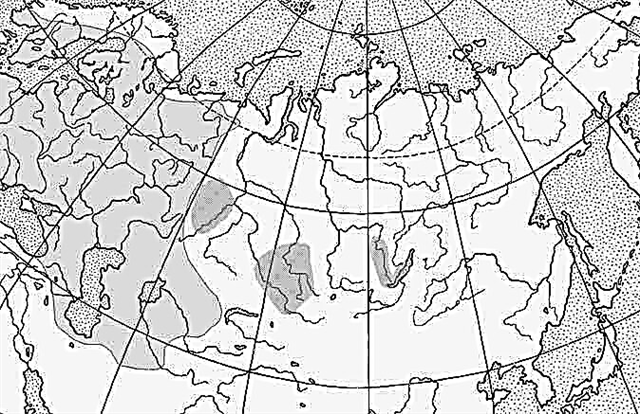
Рисунок 79. Ареал белого аиста в Вост. Европе и Сев. Азии:
а — регулярное гнездование, б — недостаточно выясненная граница гнездового ареала, в — нерегулярное гнездование. Подвиды: 1 — С. с. ciconia, 2 — С. с. asiatica.
Периодические выселения отдельных пар отмечены далеко за пределами указанной границы: в Юж. Карелии, Костромской, Нижегородской, Кировской, Пермской, Ульяновской, Пензенской, Саратовской, Волгоградской и Ростовской обл., Краснодарском крае (Лапшин, 1997, 2000, Бакка и др., 2000, Бородин, 2000, Дылюк, Галченков, 2000, Карякин, 2000, Комлев, 2000, Мнацеканов, 2000, Пискунов, Беляченко, 2000, Сотников, 2000, Фролов и др., 2000, Чернобай, 2000а и др.). Западноазиатская популяция номинативного подвида распространена в Терско-Сулакской низменности Дагестана (Бабаюртовский, Хасавюртовский, Кизлярский, Тарумовский р-ны), периодически гнёзда появлялись за пределами Дагестана — в Ставропольском крае, Карачаево-Черкесии, Пролетарском р-не Ростовской обл. (Хохлов, 1988а, Бичерев, Скиба, 1990). Регистрировали белых аистов и в предгорьях Сев. Осетии (Комаров, 1986). Ростовская обл., очевидно, является территорией, где с разных направлений практически сходятся восточноевропейская и западноазиатская популяции. Первая проникает сюда с севера по Дону и с запада — с Украины, вторая — с юго-востока по Кумо-Манычкой впадине. Подтверждением последнего, наиболее плохо выясненного направления перемещения птиц, может служить встреча 13 мая 1996 г. в районе оз. Дадинское, на крайнем северо-востоке Ставропольского края, стаи из 18 птиц, мигрировавших на большой высоте в северо-западном направлении (Дылюк, Галченков, 2000).
На Украине современная граница ареала проходит через Сев. и Сев.-Вост. Крым, южные части Запорожской и Донецкой обл., Луганскую обл. (Грищенко, 2005). В 2006 г. отмечен первый случаи гнездования белого аиста на юго-востоке Крыма возле Феодосии (М. М. Бескаравайный, личн. сообщ.).
Туркестанский белый аист распространён в Средней Азии — на юго-востоке Узбекистана, в Таджикистане, Киргизии, на юге Казахстана. Ранее ареал доходил до Чарджоу в Туркмении, низовьев Аму-Дарьи, отмечались случаи гнездования и на западе Китая — в Кашгарии (Спангенберг, 1951, Долгушин, 1960, Сагитов, 1987, Шерназаров и др., 1992). Изредка попытки гнездования — очевидно, уже европейского подвида — отмечаются на юго-востоке Туркменистана (Белоусов, 1990).
Небольшой очаг гнездования белого аиста (около 10 пар) возник на крайнем юге Африки. Птицы здесь приступают к гнездованию в сентябре-ноябре — в пору прилёта за зимовки аистов северных популяций (del Hoyo et al., 1992). Как и в случае с чёрным аистом, эта микропопуляция ведёт начало от мигрантов, по каким-то причинам начавших размножаться на зимовках.
Зимовки
Основные места зимовки западной популяции европейского подвида — саванны к югу от Сахары от Сенегала на западе до Камеруна на востоке. Важнейшие места концентрации зимующих птиц — долины рек Сенегал, Нигер и район оз. Чад. Здесь же зимуют и аисты, гнездящиеся на северо-западе Африки. Восточная популяция зимует в Вост. и Юж. Африке от Судана, Эфиопии и Сомали до ЮАР. Большая часть птиц проводит зимние месяцы в Танзании, Замбии, Зимбабве, ЮАР. Аисты из Зап. Азии зимуют частично в Африке, частично — на юге Азии. Азиатский подвид зимует в основном в Индии на юг до Шри-Ланки. На восток эти птицы могут встречаться до Таиланда (Schulz, 1988, 1998, Ash, 1989, Hancock et al., 1992). В Индии основными местами зимовки аистов служат штаты Бихар на северо-востоке и Гуджарат на западе (Majumdar, 1989). Интересно, что в Индии находили и птиц, окольцованных в Европе (Лебедева, 1979а). Видимо, это аисты, сбившиеся с пути в районе залива Искандера — не свернувшие на юг, а продолжившие миграцию на юго-восток.
Часть птиц зимует в южной части гнездового ареала. В Испании в зимние сезоны 1991 и 1992 гг. в дельте р. Гвадалквивир и на побережье Андалузии было учтено около 3 000 особей (Tortosa et al., 1995). В Португалии зимой 1994/95 гг. зимовало 1 187 аистов (Rosa et al., 1999). Тысячи аистов остаются на зиму в Израиле (Schulz, 1998). В Армении в долине Аракса ежегодно зимуют сотни птиц (Адамян, 1990). В Болгарии аисты оставались на зимовку ещё в конце XIX в., сейчас их количество значительно возросло. Отмечаются стаи до 10 особей (Nankinov, 1994). Известны случаи зимовки и в более северных широтах — на Украине (Грищенко, 1992), в Чехии (Tichy, 1996), Германии, Дании (Schulz, 1998). На территории России зимовки белых аистов отмечались в Дагестане (Т.К. Умаханова, В. Ф. Маматаева, личн. сообщ.). В Средней Азии аисты зимуют в небольшом количестве в Ферганской долине (Третьяков, 1974, 1990). Здесь в районе Пунгана — Ургенча в зимние месяцы 1989 г. регистрировали до 250 птиц. Полагают, что частичная оседлость белых аистов в Ферганской долине способствовала общему увеличению их количества в регионе. Зимовки, носящие нерегулярный характер, наблюдали в долине Сыр-Дарьи и на р. Пяндж в Юж. Таджикистане (Митропольский, 2007).
Окольцованные в бывш. СССР белые аисты обнаружены на зимовке в основном в ЮАР, отдельные птицы — в Эфиопии, Судане, Уганде, Кении, Намибии, Зап. Африке (Лебедева, 1979, Смогоржевський, 1979).
Как установил X. Шульц (Schulz, 1988), распределение аистов на местах зимовки в Африке определяется прежде всего запасами корма. В первую очередь птицы выбирают влажные биотопы, но могут держаться и в богатых кормом засушливых местах. Большие стаи встречаются даже в пустынях и горах. В Лесото в 1987 г. была обнаружена стая из 200 аистов на высоте ок. 2 000 м. Птицы кормились на водоёмах, изобиловавших земноводными. В местах, богатых кормом, аисты могут скапливаться в огромных количествах. В январе 1987 г. в Танзании на участке в 25 км2 было учтено-около 100 тыс. особей. Птицы кормились на люцерновых полях, где массово размножились гусеницы одной из местных бабочек. В Юж. Африке в этот сезон белые аисты почти не встречались.
Благодаря результатам кольцевания и спутниковой телеметрии установлено, что места зимовки западной и восточной популяций не изолированы друг от друга. В Центр. Африке существует зона смешанных зимовок, где встречаются птицы из обеих популяций. Здесь особи из одной популяции могут увлекаться стаями аистов из другой популяции и возвращаться весной другим путем и в другие места гнездования (Berthold et al., 1997, Brouwer et al., 2003).
Миграции
Белый аист — дальний мигрант. Птицы из северо-восточной части ареала совершают перелеты более 10 тыс. км. Существуют две основные географические популяции европейского подвида, различающиеся путями пролёта и местами зимовки. Линия раздела между ними проходит через Голландию, Гарц, Баварию, Альпы (Schuz, 1953, 1962, Creutz, 1988, Schulz, 1988, 1998). Птицы, гнездящиеся к западу от неё, мигрируют осенью на юго-запад через Францию, Испанию, Гибралтар. Далее пролёт идёт через Марокко, Мавританию, западную часть Сахары. Зимуют эти птицы в Зап. Африке. Аисты, гнездящиеся к востоку от этой разделительной линии, летят осенью в юго-восточном, а из России, Украины, Белоруссии и Прибалтики — в южном направлениях. Через территорию Украины осенью проходит три основных пролётных пути, которые сливаются в мощный миграционный поток, проходящий по западному побережью Чёрного моря (Грищенко, Серебряков, 1992, Grischtschenko et al., 1995). Далее аисты летят через Балканы и Турцию, через Босфор, Малую Азию. От Искандера они выходят к средиземноморскому побережью, где снова поворачивают на юг и узким потоком мигрируют через Ливан, Израиль, Синайский п-ов к долине Нила. Вдоль этой реки и Рифтовой долины идёт дальнейшая миграция к основным местам зимовки в Вост. и Юж. Африке. В Вост. Судане аисты делают длительную остановку на 4-6 недель и интенсивно кормятся, чтобы восстановить жировые резервы для продолжения миграции (Schulz, 1988, 1998).
Аист, как сухопутный паритель, избегает длительного перелёта над морем, поэтому образуются миграционные потоки вдоль побережий. Аисты из западных, северных и центральных областей Украины мигрируют вдоль западного побережья Чёрного моря и через Босфор, а птицы из Вост. Украины летят на юго-восток к восточному побережью Чёрного моря. Сюда же летят и аисты из восточной части ареала в России. Часть аистов, хотя и незначительная, все же летит напрямую через море. Существует «промежуточный» пролётный путь через Италию и Сицилию в Тунис. В 1990-1992 гг. на мысе Бон в Тунисе учтено 1 378 мигрировавших аистов и 67 — возле Мессины в Сицилии (Kisling, Horst, 1999). Предполагается, что этот путь используют птицы как из западной, так и из восточной популяций (Schulz, 1998). Особь, окольцованная в Латвии, обнаружена в сентябре у Неаполя (Лебедева, 1979). А один аист со спутниковым передатчиком перелетел через Средиземное море напрямую от г. Сан-Тропе во Франции до Туниса, путь через море составил как минимум 752 км (Chemetsov et al., 2005). Возможно, часть аистов перелетает через Чёрное море, пересекая Крым.
Миграции аистов из Закавказья, Ирака и Ирана изучены недостаточно. Предполагается, что они улетают на юго-восток в Юж. Азию (Schtiz, 1963, Schulz, 1998). Птица, окольцованная в Армении, была найдена в Нахичеванской АССР в 160 км к юго-востоку (Лебедева, 1979). Линия раздела между популяциями, мигрирующими в Африку и в Азию пока неизвестна. Видимо, она проходит где-то на востоке Турции. По крайней мере, в этом регионе осенью наблюдают стаи птиц, мигрирующих как на юго-восток, так и на запад (Schtiz, 1963).
Туркестанские аисты осенью улетают на юг через Афганистан в Индию, пересекая Гиндукуш через перевал Саланг (Schtiz, 1963, Schulz, 1998). Окольцованных в Узбекистане аистов добывали весной в Афганистане и Пакистане (Лебедева, 1979).
Анализ спутникового прослеживания 140 немецких аистов показал, что у этих птиц могут в довольно широких пределах изменяться пути и сроки миграции, места зимовки и остановок, но, при возможности, они остаются постоянными. Изменения вызываются природными факторами, прежде всего условиями кормёжки (Berthold et al., 2004). Сроки отлёта с мест зимовки зависят от метеорологической ситуации. При неблагоприятных условиях птицы могут задерживаться. Так, в крайне неблагоприятном 1997 г. аисты стартовали с мест зимовки на месяц позже обычных сроков (Косарев, 2006). К этому добавилась задержка из-за длительных холодов на Ближнем Востоке. Снабжённые передатчиками аисты совершали длительные остановки в Сирии и Турции. Отмечался обратный пролёт (Kaatz, 1999). В итоге в 1997 г. только 20% птиц из восточной популяции прилетели в нормальные сроки, большинство же — с задержкой в 4-6 недель (Schulz, 1998).
С мест зимовки массовое движение в обратном направлении происходит в конце января или в феврале. В Израиле начало весенней миграции взрослых птиц становится заметным в середине февраля, пик пролёта приходится на вторую половину марта, заканчиваются особенно заметные перемещения в конце апреля, молодые птицы мигрируют через Израиль в апреле-мае (van den Bossche et al., 2002). В местах гнездования на севере Африки аисты появляются ещё в декабре-феврале. Пик пролёта над Гибралтаром отмечают в феврале-марте, над Босфором — с конца марта до конца апреля (Schulz, 1998).
В Молдавии прилетевшие аисты отмечаются с первой декады марта (Аверин и др., 1971). На территории Украины прилёт регистрируется с первых чисел марта до второй половины апреля, средние даты прилёта приходятся на третью декаду марта — начало апреля. Раньше всего птицы появляются во Львовской и Черновицкой обл., облетая Карпаты, затем миграция идёт двумя потоками: одни птицы летят на северо-восток, другие — на восток по южным областям Украины. Позже всего аисты появляются в восточных областях и в Крыму (Грищенко, Серебряков, 1992, Grischtschenko et al., 1995). На севере Сумской обл. прилёт регистрировали с 18 марта по 26 апреля, средняя дата за 16 лет — 30 марта (Афанасьев, 1998). На юго-западе Белоруссии прилёт аистов отмечается в третьей декаде марта — первой половине апреля (Шокало, Шокало, 1992). Аисты, гнездящиеся в европейской части России, достигают родины в начале марта -первой половине мая. На территории Калиниградской обл. в первой половине XX в. первые птицы появлялись на гнёздах в период с 19 марта по 12 апреля (данные за 23 года, Tischler, 1941). В 1970-е гг. прилёт аистов происходил с начала марта (Беляков, Яковчик, 1980). В 1990 г. первые птицы на гнёздах в Калининградской обл. отмечены 18 марта (Гришанов, Савчук, 1992). В Себежском р-не Псковской обл. прилёт наблюдали в конце марта-первой декаде апреля (Фетисов и др., 1986). За период с 1989 по 1999 гг. наиболее ранняя регистрация в Калужской обл. зафиксирована 20 марта (1990 г.), наиболее поздняя — 8 апреля (1991 и 1997 гг.), в среднем 30 марта. В отдельные годы самые ранние птицы появляются весной при высоте снегового покрова на полях 30-40 см. Пик прилёта первых аистов на гнёзда в Калужской обл. приходится на вторую пятидневку апреля (1990-1999 гг.) (Галченков, 2000). В Воронежской обл. первых аистов наблюдали в те же сроки: с 19 марта по 8 апреля, в среднем 30 марта (1995-1998 гг.) (Нумеров, Макагонова, 2000). К северо-восточной границе ареала аисты прилетают на 2-4 недели позже. В Ярославскую обл. птицы прилетали 22-26 апреля (1994 г.), 16 апреля (1996 г.), 2 мая (1995 г.) (Голубев, 2000). В восточных районах Лениградской обл. самый ранний прилёт отмечен 20 апреля 1999 г. (Тихвинский р-н), обычные же сроки — с 1 по 8 мая (1983-1999 гг.) (Храбрый, 2000). В южных районах Карелии первые птицы появляются в конце апреля — середине мая, в очень раннюю весну 1990 г. одиночную птицу видели в начале второй декады апреля (Лапшин, 2000). В Кировской обл. наиболее ранняя регистрация белого аиста — 17 апреля 1992 г. (Сотников, 2000). На черноморском побережье Сев. Кавказа весенний пролёт отмечается с первой декады марта до второй половины апреля, в Ростовской обл. и Краснодарском крае первых птиц отмечали в апреле (Казаков и др., 2004). В Дагестане первые особи появляются в начале и середине марта (Маматаева, Умаханова, 2000).
Появление белых аистов весной в Средней Азии приходится на конец февраля — начало марта и на большей части территории отмечается практически одновременно (Дементьев, 1952, Митропольский, 2007). На перевале Чокпак их регистрировали 11-14 марта 1974 г. (Гаврилов, Гисцов, 1985), интенсивный пролёт отмечен 24 марта (Сема, 1989).
В Калужской обл. в 69% случаев прилёт белого аиста проходил по схеме 1+1: сначала прилетала одна птица из пары, через некоторое время после неё — вторая. Первая особь появляется с 20 марта по 18 мая, в среднем (п = 176) — 10 апреля, вторая — с 25 марта по 26 мая, в среднем (п = 150) — 14 апреля. Запаздывание второй птицы происходит в интервале от нескольких часов до 31 дня, в среднем, на 4 дня. В указанной схеме прилёта существуют редко встречающиеся варианты: при первом каждая из особей пары подлетает с одной или двумя другими птицами, которые на гнезде не остаются, а летят дальше, при втором — к одиночному аисту прилетает пара и изгоняет его. В 31% случаев на гнездо прилетали сразу две птицы.
Гнездящиеся птицы восточноевропейской популяции улетают в августе. Молодые, как правило, отлетают раньше взрослых птиц. В Калужской обл. молодые покидали гнёзда начиная с 8 августа, чаще во второй декаде этого месяца. Взрослые птицы оставляют родину позже, отлёт последних особей заканчивается в среднем 30 августа (1985-1999 гг.) (Галченков, 2000). В Тверской обл. аисты отлетают 28 августа — 5 сентября (Николаев, 2000). В Ярославской обл. птицы отлетали 23 августа (1996 г.) и 29 августа (1995 г.) (Голубев, 2000). Отдельные особи и пары задерживаются до сентября — октября. В юго-западных регионах России перед отлётом образуют скопления в десятки и до 100 и более особей, как например, в Смоленской обл. (Бичерев, Барнев, 1998). На Сев. Кавказе осенняя миграция отмечается с первой половины августа до конца сентября (Казаков и др., 2004). Не выяснены пути миграции и районы зимовки дагестанских аистов: известно, что гнездовую область последние из них покидают с 25 октября по 10 ноября, иногда задерживаясь до середины-конца этого месяца (25 ноября 2003 г. и 15 ноября 2004) г. Вероятнее всего аисты, гнездящиеся в Терско-Сунженской низменности, следуют вдоль западного берега Каспийского моря, где птицы этого вида были отмечены 23 октября 1998 г. в районе г. Каспийск (Е.В. Вилков, личн. сообщ.).
В Молдавии отлёт начинается в конце августа и продолжается до середины сентября. Отдельные птицы могут задерживаться до первой половины октября. Наиболее поздняя встреча — 9 ноября 1964 г. (Аверин, Ганя, Успенский, 1971). На Украине первые пролётные стаи наблюдаются с первой декады августа до сентября и начала октября. Средние даты начала отлёта приходятся на третью декаду августа — первую декаду сентября. Раньше всего пролёт начинается во Львовской, Житомирской и Полтавской обл. Последних птиц наблюдали со второй половины августа до октября. Средние даты последнего наблюдения в большинстве областей Украины приходятся на первую и вторую декады сентября. Дольше всего аисты задерживаются в Запорожской обл. и в Крыму (Грищенко, Серебряков, 1992, Grischtschenko et al., 1995). Отдельные запоздавшие особи могут отмечаться и в ноябре. Иногда очень поздно можно встретить целые стаи. Так, 4 декабря 1985 г. над Ивано-Франковском наблюдали стаю из нескольких десятков аистов (Штыркало, 1990). 5 ноября 1997 г. над Брестом видели стаю из 40 особей (Шокало, Шокало, 1992). Пролёт по восточному побережью Чёрного моря отмечали с 29 августа по 4 октября (Абуладзе, Элигулашвили, 1986).
Среднеазиатские аисты отлетают с конца августа до средины октября (Долгушин, 1960, Третьяков, 1990).
Перелёт трёх молодых аистов, помеченных на гнёздах в Зеленоградском и Гурьевском р-нах Калининградской обл. спутниковыми передатчиками, был прослежен в 2000 г. Одна птица отправилась на зимовку 10 августа, две другие — 14-го. Пролётный путь прошёл через северо-восточную Польшу, крайний юго-запад Белоруссии, западную часть Украины, восток Румынии и Болгарии, далее через пролив Босфор, Турцию, Палестину, Синайский п-ов. Пролива Босфор аисты достигли, соответственно, 23, 25 и 26 августа, т.е. через 13, 11 и 12 дней после начала миграции. На южной оконечности Синайского п-ова аисты были 29, 31 августа и 1 сентября соответственно (через 19, 17 и 18 дней после начала миграции, или через 6 дней после пересечения Босфора каждой из птиц), здесь аисты сделали остановку. Далее аисты проследовали вдоль долины Нила в континентальном Египте. Быстрое движение к югу птицы прекратили 6, 7 и 10 сентября, к этому моменту две из них находились в центр. Судане, одна — на востоке Чада поблизости от суданской границы (Chemetsov et al., 2004).
В ходе осенних миграций средняя протяжённость суточных перемещений аистов восточной популяции по данным телеметрии составляет: в Европе — 218 км (для взрослых птиц от 52 до 504, для молодых — от 51 до 475 км), на Ближнем Востоке — 275 км (для взрослых птиц от 52 до 490, для молодых — от 55 до 408 км), в Сев. Африке — 288 км (для взрослых птиц от 70 до 503, для молодых — от 108 до 403 км) (van den Bossche et al., 1999).
Комплексное исследование миграций белого аиста показало, что у этого вида, по крайней мере его восточной популяции, совершенно особый тип миграции, пока не известный для других птиц. Для него характерен очень быстрый перелёт от мест гнездования к зоне отдыха в Вост. Африке. Расстояние в 4 600 км, как взрослые, так и молодые птицы преодолевают в среднем за 18-19 дней. При нормальных условиях аисты летят ежедневно, проводя в пути по 8-10 ч. Длительные, особенно многодневные остановки, встречаются лишь как исключение и связаны прежде всего с неблагоприятными погодными условиями. У аистов, в отличие от других перелётных птиц, жировые запасы во время миграции незначительны. Заметной гиперфагии во время перелёта не наблюдается. Аисты практически не восстанавливают вес до самой Африки (Berthold et al., 2001).
Большинство неполовозрелых аистов проводят летние месяцы вдали от мест гнездования. После первой зимовки птицы мигрируют в направлении области гнездования, но достигают её очень редко. Только треть однолетних аистов была обнаружена ближе 1000 км от места кольцевания. С возрастом доля «невозвращенцев» быстро уменьшается. Значительная часть 1-2-годовалых аистов проводят лето южнее Сахары, но 3-летние птицы в гнездовой период там не встречаются вообще. Кольцевание показало, что в большинстве случаев аисты впервые появляются в местах гнездования в 3-летнем возрасте (Libbert, 1954, Kania, 1985, Bairlein, 1992).
Залётные аисты могут встречаться значительно севернее и восточнее границы гнездового ареала. В России они отмечались на берегу Белого моря в Мурманской обл. (Коханов, 1987), у с. Холмогоры в Архангельской обл. (Плешак, 1987), в Башкирии (Карякин, 1998а), Татарстане (Аськеев, Аськеев, 1999), Пермской обл. (Демидова, 1997, Карякин, 19986), Свердловской обл. (Зеленцов, 1995), в степях Юж. Урала (Давыгора, 2006). По недостаточно надёжным данным, двух птиц наблюдали в августе в Курганской обл. (Тарасов и др., 2003). Залёты белых аистов регистрируют также в Финляндии, Швеции, Норвегии, Великобритании, Ирландии, Исландии (Hancock et al., 1992, Бирина, 2003). Во время миграции могут отмечаться настоящие инвазии, когда большие стаи оказываются вдали от основных пролётных путей. Так, 15 сентября 1984 г. возле г. Абу-Даби на востоке Аравийского п-ова появилась стая из 3 000 аистов (Reza Khan, 1989). 27-29 августа 2000 г. скопление из 300-400 особей держалось в долине р. Теберда на Сев. Кавказе (Поливанов и др., 2001). Иногда пролётные стаи аистов заносит ветром далеко в море. Такие птицы отмечаются даже на Сейшельских о-вах, лежащих за 1 000 км от африканского побережья (Stork, 1999).
Местообитание
Белый аист — типичный обитатель открытых ландшафтов, сплошных лесов и заросших болот избегает. Предпочитает территории с влажными биотопами — лугами, болотами, пастбищами, поливными землями, рисовыми полями и т.п. Встречается также в степях и саваннах с одиночными крупными деревьями или сооружениями человека. Оптимальным биотопом в наших условиях являются обширные поймы рек с нормальным гидрорежимом и экстенсивным сельскохозяйственным использованием. В таких местах плотность населения может достигать десятков пар на 100 км2. Населяет, как правило, равнинные территории, но может гнездиться и невысоко в горах с подходящими условиями.
В Центр. Европе белые аисты редко гнездятся выше 500 м н. у.м (Schulz, 1998). В Карпатах они поднимаются до 700-900 м (Смогоржевський, 1979, Rejman, 1989, Stollmann, 1989), в Армении и Грузии — до 2 000 м н.у.м. (Адамян, 1990, Gavashelishvili, 1999), в Турции до 2 300 м (Creutz, 1988), а в Марокко даже до 2 500 м н.у.м (Sauter, Schiiz, 1954). В Болгарии 78,8% аистиных пар гнездится на высотах от 50 до 499 м н.у.м. и только 0,2% — от 1 000 до 1 300 м (Petrov et al., 1999). В Польше отмечено расселение аистов на более значительные высоты в ходе роста численности (Tryjanowski et al., 2005). Кормиться белый аист предпочитает на открытых участках с невысокой травянистой растительностью, на мелководье стоячих и медленно-проточных водоёмов. Реже встречается на берегах крупных рек, горных ручьёв. Пахотные земли и интенсивно культивируемые луга и поля многолетних трав также используются аистами для кормёжки, однако благоприятный период для сбора корма в таких местах очень короткий — сразу после вспашки или сбора урожая.
Гнёзда аиста встречаются на периферии колоний цапель и других голенастых птиц. Но чаще всего он гнездится в населённых пунктах. Может селиться даже среди плотной застройки в больших городах, откуда ему приходится летать за кормом за 2-3 км. Покинутые людьми села белый аист обычно со временем оставляет. Так, перестали эти птицы гнездиться в большинстве выселенных сёл Чернобыльской зоны (Samusenko, 2000, Гашек, 2002).
Во время миграции белый аист также предпочитает открытый ландшафт, большие водные пространства и лесные массивы старается облетать, поскольку, как мы считаем, полет над ними у специализированного парителя требует большего расхода энергии.
Численность
Общую численность белого аиста по результатам V Международного учёта в 1994-1995 гг. можно оценить минимум в 170-180 тыс. пар, из них на восточную популяцию приходится 140-150 тыс. пар (Грищенко, 2000). По сравнению с предыдущей переписью в 1984 г., общая численность выросла на 23%. Причём численность западной популяции увеличилась значительно больше — на 75%, восточной — на 15% (Schulz, 1999). Наибольшее количество белого аиста отмечено в Польше. В 1995 г. там учтено около 40900 пар, на 34% больше, чем в 1984 г. Средняя плотность гнездования в Польше составляет 13,1 пар/100 км2 (Guziak, Jakubiec, 1999). В Испании, где гнездится основная часть западной популяции, в 1996 г. численность оценивалась в 18 000 пар. В этой стране отмечен наибольший её рост: между двумя международными переписями она увеличилась здесь более, чем вдвое (Marti, 1999).
По предварительным итогам VI Международного учёта, проходившего в 2004-2005 гг., общая численность белого аиста оценивается в 230 тыс. пар. Наибольшая численность в Польше — 52,5 тыс. пар, далее идут Испания — 33,2 тыс. пар, Украина — ок. 30 тыс. пар, Белоруссия — 20,3 тыс. пар, Литва — 13 тыс. пар, Латвия — 10,7 тыс. пар, Россия — 10,2 тыс. пар. Наибольший прирост численности отмечен во Франции — 209%, Швеции — 164%, Португалии— 133%, Италии— 117%, Испании— 100%. Снизилась (вдвое) численность только в Дании. Там осталось всего 3 гнезда. По азиатскому подвиду данные приводятся только для Узбекистана, где учтено 745 пар, численность снизилась на 49%.
В ходе VI Международного учёта, по предварительным данным, учтено: Курская обл. — 929 пар (+186% по сравнению с V Международным учётом, данные В. И. Миронова), Брянская обл. — 844 (+31%, С. М. Косенко), Калужская обл. — 285 (+58%, Ю. Д. Галчёнков), Ленинградская обл. — 160 (+344%, В.Г. Пчелинцев), Орловская обл. — 129 (С. В. Недосекин), Московская обл. — 80 (+248%, М. В. Калякин).
Современная численность в Армении оценивается в 1-1,5 тыс. пар, в Азербайджане — 1-5 тыс. пар, в Молдавии — 400-600 пар (Birds in Europe, 2004).
На протяжении XX столетия численность белого аиста подвергалась значительным изменениям (см. Грищенко, 2000). В первой половине века (а местами ещё раньше) началось быстрое её сокращение во многих странах Европы. К концу 1940-х гг. в Средней Европе она уменьшилась почти вдвое. Проводившиеся в 1934, 1958, 1974, 1984 гг. международные учёты белого аиста показывали неуклонное снижение числа занятых гнёзд. Так, если в 1907 г. в Германии насчитывалось 7-8 тыс. гнездящихся пар (Wassmann, 1984), то к 1984 г. их количество уменьшилось до 649 в ФРГ (Heckenroth, 1986) и 2 724 в ГДР (Creutz, 1985). В Нидерландах в XIX в. белый аист был одной из обычных птиц, в стране были тысячи гнёзд. Но уже в 1910 г. оставалось всего 500 гнездящихся пар, численность продолжала стремительно уменьшаться: 209 пар в 1929 г., 85 — в 1950 г., 5 — в 1985 г. (Jonkers, 1989). После 1991 г. не осталось вообще ни одной «дикой» пары, гнездились только птицы, выпущенные из специальных питомников (Vos, 1995). Перестали аисты гнездиться в Бельгии, Швейцарии, Швеции, подошли к грани исчезновения во Франции, Дании и некоторых других странах. Наиболее уязвимой оказалась западная популяция белого аиста. По данным IV Международного учёта в 1984 г., только за 10 лет численность её сократилась на 20%, восточной популяции — на 12% (Rheinwald, 1989).
Радикальное изменение ситуации началось в 1980-х гг., раньше всего в Испании. Примерно с 1987 г. начинается рост количества аистов. За 11 лет оно увеличилось более чем в 2,5 раза и вскоре превысило уровень полувековой давности (Gomez Manzaneque, 1992, Martinez Rodriguez, 1995). Более чем в 2 раза выросла численность и в Португалии (Rosa et al., 1999). Все это объяснялось прежде всего климатическими причинами. Во второй половине 1980-х гг. наконец прекратился длительный период засухи в зоне Сахеля, значительно ухудшивший условия зимовки западной популяции белого аиста. Способствовало росту численности и значительное улучшение кормовой базы на местах гнездования. В Испании, например, увеличилась площадь орошаемых земель, кроме того в каналах прижился южноамериканский рак Procambarus clarkii, которого охотно поедают аисты (Schulz, 1994, 1999). В Испании и Португалии гораздо больше птиц стало оставаться на зимовку, что также снизило смертность (Gomez Manzaneque, 1992, Rosa et al., 1999). Скачок численности белого аиста на Пиренейском п-ове способствовал быстрому росту всей западной популяции. Вскоре подъем численности и расселение этих птиц начались во Франции, причём связь с процессами, происходившими в Испании, была доказана: в 1990 и 1991 гг. нашли аистов, гнездившихся на Атлантическом побережье Франции, а окольцованных в Испании. Предполагается, что часть аистов, загнездившихся в департаментах по побережью Бискайского залива, расселилась из Испании. На северо-востоке и в центре Франции появились аисты из Эльзаса, Швейцарии и Нидерландов. В департаменте Приморская Шаранта в 1995 г. гнездился аист, окольцованный в 1986 г. птенцом в Польше. Отмечено также быстрое расселение аистов в Голландии, Швейцарии, Италии, Германии и других странах. Во Франции с 1984 по 1995 гг. численность увеличилась на 830% (Duquet, 1999).
У восточной популяции не было таких резких скачков численности, как у западной, но отмечен её положительный тренд. Подчеркнём, что при общем снижении численности продолжалось расселение аистов на восток в России и на Украине и её рост у границы ареала. Увеличение численности восточной популяции началось примерно в те же сроки, что и западной, хотя темпы роста были гораздо ниже. Практически синхронно изменилась ситуация и у азиатского подвида. С 1984 до 1994 г. численность белого аиста в Средней Азии выросла более чем в 7 раз (Shemazarov, 1999), а к 2005 г. количество этих птиц здесь оценивают в 700-1 000 гнездящихся пар (Митропольский, 2007).
По данным мониторинга на постоянных пробных участках на Украине, в 1990 гг. прошла волна роста численности. Она наметилась уже в первой половине 1990-х гг., несколько раньше на северо-востоке Украины, позже всего — в западных областях. В 1992-1994 гг. в сёлах по р. Сейм в Сумской обл. отмечалось увеличение численности на 25-30% ежегодно (Грищенко, 1995а, 20006). С 1994 г. средний по Украине прирост все время увеличивался (спад отмечен лишь в 1997 г., крайне неблагоприятном для белого аиста по всей Европе), достигнув максимума в 1996 и 1998 гг. — соответственно 13,7±2,9 и 16,3±3,6 %. Затем темпы роста начали уменьшаться, а в 2001-2003 гг. популяция стабилизировалась. (Грищенко, 2004).
В этот же период активизировалось расселение на восток в восточных областях Украины и в России. В Харьковской обл. к 1994 г. отмечено смещение границы ареала на восток по сравнению с распространением в 1974-1987 гг., в 1998 г. обнаружены гнёзда на правом берегу р. Оскол (Атемасова, Атемасов, 2003). В Луганской обл., где белый аист встречался на восток до р. Айдар, в 1998 г. найдены 2 гнезда в пойме р. Деркул на границе с Россией (Ветров, 1998). В Ростовской обл. в 1996 г. аисты снова загнездились после 5-летнего перерыва — было обнаружено гнездо в долине Маныча (Казаков и др., 1997). В Краснодарском крае аисты начали гнездиться в середине 1990-х гг. (Мнацеканов, 2000). В 1993 г. гнездование впервые отмечено в Кировской обл. (Сотников, 1997, 1998), в 1994 г. — в Тамбовской обл. (Евдокишин, 1999), в 1995 г. — в Мордовии (Лапшин, Лысенков, 1997,2000), в 1996 г. — в Вологодской обл. (Дылюк, 2000). В 1996 г. отметили резкое увеличение количества птиц (на 20,1%) в Калужской обл. (Галченков, 2000).
Суточная активность, поведение
Белый аист — дневная птица, однако известны случаи кормления птенцов и в светлые ночи (Schuz, Schuz, 1932). Ночью птицы могут проявлять активность на гнезде: отмечались копуляции, уход за оперением, смена насиживающих партнёров и т. п. Во время миграции аист летит днём, но на северо-западе Африки при высокой дневной температуре пролётные стаи отмечались и ночью (Bauer, Glutz von Blotzheim, 1966). Большие стаи чаще всего скученные, неупорядоченные, птицы летят на разной высоте (Молодовский, 2001).
По земле белый аист передвигается шагами, реже бегает. Активный полёт довольно тяжёлый, с медленными взмахами крыльев. При благоприятных условиях предпочитает парение, особенно при полёте на большие расстояния. В восходящих потоках нередко образуются скопления набирающих высоту птиц. Белый аист может плавать, хотя делает это неохотно. При благоприятном ветре способен взлетать с поверхности воды (Bauer, Glutz von Blotzheim, 1966, Creutz, 1988).
Во внегнездовой период белый аист ведёт стайный образ жизни. Во время гнездования также могут образовываться колонии и скопления в местах кормёжки. Неразмножающиеся птицы летом держатся стаями, численность которых достигает десятков и даже сотен особей. Они держатся в богатых кормом местах, ведя бродячий образ жизни. Численность таких стай постепенно увеличивается от мая к июню, в июле их размеры заметно возрастают, ещё более многочисленными они становятся в августе, в связи с образованием предотлётных скоплений. По наблюдениям в Калужской обл. в 1990-е гг., средняя численность птиц в летних стаях составляла: в мае — 3,4 особи, в июне — 4,0, в июле — 7,8, в августе — 10,5 (п = 50). Выводки после вылета объединяются в стаи, которые постепенно укрупняются по ходу миграции. Так, если на Украине обычный размер пролётных стай осенью — десятки, реже сотни особей, то уже на черноморском побережье Болгарии средний размер стаи составляет 577,5 особи (Michev, Profirov, 1989). На Ближнем Востоке и в Сев.-Вост. Африке нередко отмечают огромные скопления, превышающие 100 тыс. особей (Schulz, 1988, 1998). Установлено, что эффективность миграции (скорость передвижения, компенсация сноса ветром и т.п.) выше в больших стаях (численностью в несколько тысяч особей), чем в мелких группах или у отдельных птиц (Liechti et al., 1996).
Отдыхают аисты преимущественно ночью. В гнездовой период количество времени, которое остаётся для отдыха и чистки оперения, зависит от обилия пищи и количества птенцов. При её изобилии, аисты могут и днём часами отдыхать или чистить оперение. Поза отдыхающей птицы весьма характерна: аист стоит чаще всего на одной ноге, втянув голову в плечи и пряча клюв в распушённое оперение шеи. Как правило, отдыхают аисты на высоких присадах с хорошим обзором — на сухих деревьях, столбах, крышах.
Белые аисты используют довольно необычный способ терморегуляции — испражняются на ноги. В жаркий день можно увидеть многих птиц с белыми «чулками» на лапах. По-видимому, жидкая мочевая кислота испаряется, охлаждая поверхность цевки. Кожа её обильно пронизана кровеносными сосудами, проходя через которые кровь охлаждается (Prinzinger, Hund, 1982, Schulz, 1987). Как показали эксперименты на американском лесном аисте (Mycteria americana), при интенсивном испражнении на ноги температура тела снижается (Kahl, 1972). X. Шульц (Schulz, 1987), наблюдая за аистами в Африке, установил, что частота испражнений зависит от температуры воздуха. Температурный порог, после которого птицы с забрызганными помётом ногами начинают встречаться регулярно, равен примерно 28 °С. При 40° частота испражнений доходит уже до 1,5 раз в минуту. Белый помёт, кроме того, экранирует ноги от палящих лучей солнца. При облачной погоде частота испражнений снижается. Наблюдения на Украине показали, что в местах гнездования аисты также начинают использовать такой способ терморегуляции при температуре около 30 °С (Grischtschenko, 1992).
При совместной кормёжке белого и чёрного аистов и цапель доминирует белый аист (Kozulin, 1996).
Питание
Питание белого аиста очень разнообразно. Он поедает различных мелких животных от дождевых червей до грызунов и мелких птиц: пиявок, моллюсков, пауков, ракообразных, насекомых и их личинок, рыб, земноводных, пресмыкающихся и т.п. Может разорять гнёзда наземногнездящихся птиц или поймать зайчонка. В питании отмечены даже мелкие хищники, такие как ласка (.Mustela nivalis) (Lohmer et al, 1980, Штыркало, 1990). Размер добычи ограничивается лишь возможностью её проглотить. Рацион зависит от условий местности и численности объектов охоты. В сухих местах он почти полностью может состоять из насекомых, на лугах и болотах их доля значительно меньше. Так, по данным Э. Г. Самусенко (1994), в Белоруссии удельный вес различных групп животных в питании белого аиста колеблется в значительных пределах. В поймах Сожа и Березины беспозвоночные составляли по частоте встреч 51,6-56,8%, во внепойменных биотопах — до 99%.
Добычу аисты глотают целиком. Мелких животных проглатывают сразу, крупных насекомых и грызунов сначала убивают ударами клюва. Иногда можно видеть, как белый аист некоторое время как бы «жуёт» клювом пойманную полёвку или крота. Крупную сухую добычу при наличии поблизости воды птица некоторое время прополаскивает, пока её можно будет легко проглотить. Таким же образом аисты отмывают запачканных в иле лягушек или pbi6y(Creutz, 1988).
Непереваренные остатки пищи отрыгиваются в виде погадок. Погадки формируются на протяжении 36-48 ч. Состоят они из хитиновых остатков насекомых, шерсти и костей млекопитающих, чешуй рыб и пресмыкающихся, щетинок червей и т.п. Размер погадок 20-100×20—60 мм, вес 16–65 г. У птенцов они несколько мельче — 20—45×20—25 мм (Creutz, 1988, Muzinic, Rasajski, 1992, Schulz, 1998).
Кормятся аисты в разнообразных открытых биотопах — на лугах, пастбищах, болотах, берегах водоёмов, полях, огородах и т.п. Излюбленные места кормёжки — участки с нарушенной растительностью или почвенным слоем, где лишённые убежищ мелкие животные становятся лёгкой добычей. Эффективность охоты в таких ситуациях может быть весьма значительной. Так, в Польше кормившийся за убиравшим пшеницу комбайном аист за 84 мин поймал 33 грызуна (Pinowski et al., 1991). По наблюдениям в пойме Эльбы в Германии, наибольшая эффективность охоты (в среднем 5 г добычи в минуту) была во время сенокошения или сразу после него (Dziewiaty, 1992). Поэтому скопления кормящихся аистов можно увидеть на свежих покосах, на обрабатываемых полях и даже среди горящей травы. В Африке эти птицы собираются там, где местные жители выжигают саванну в сухой сезон. Достаточно им увидеть дым, как аисты слетаются отовсюду, концентрируясь за стеной огня. Они вышагивают по ещё дымящимся стеблям и вылавливают насекомых. Иногда на таких пожарищах собираются сотни особей (Creutz, 1988). Охотно аисты сопровождают стада домашнего скота или диких животных на пастбищах. Копытные вспугивают мелких животных, облегчая их добычу. На лугах аисты чаще всего кормятся на участках с низкой травой или на мелководье водоёмов. Глубже 20-30 см забредают редко. Дождевых червей аисты собирают чаще всего после дождей, когда те вылезают на поверхность, или на свежевспаханных полях. Охотно кормятся на поливных полях, изобилующих дождевыми червями. Хотя в высокой растительности численность насекомых выше, эффективность охоты белого аиста при этом снижается. Так, в Австрии она составляла 61% в растительности высотой 25 см и 52% при высоте растений 25-30 см (Schulz, 1998).
Основной способ охоты белого аиста — активный поиск добычи. Птица размеренно вышагивает по траве или мелководью, то замедляя, то ускоряя темп, может делать резкие броски или замирать на месте. Реже аисты подкарауливают жертву, прежде всего грызунов и крупных насекомых. Корм птицы собирают на земле, на мелководье, реже — на растениях. Могут они ловить клювом и пролетающих животных — стрекоз, жуков и других насекомых. Иногда даже сбивают их крыльями. Содержащиеся в неволе аисты быстро обучаются хватать на лету клювом брошенную им пищу. Описаны даже случаи успешной охоты аистов на пролетающих мимо воробьёв и других мелких птиц (Niethammer, 1967, Creutz, 1988, Berthold, 2004). Дождевых червей и других почвенных беспозвоночных птица нащупывает клювом, погружая его в землю на несколько сантиметров (Schulz, 1998). Наблюдали также, что аисты в полете схватывали рыбу с поверхности воды (Neuschulz, 1981, Schulz, 1998).
По исследованиям П. Закла (Sackl, 1985, цит. по: Schulz, 1998) в Австрии, средняя скорость передвижения аиста во время кормёжки — 1,7 км/ч. При этом он делает от 1 до 90 шагов в минуту, в среднем 39,3. Время подкарауливания добычи варьирует от 10,5 до 720 сек, в среднем составляя 151,8 сек. Изредка птицы могут замирать на месте до 12 и даже 20 мин. Кормящийся аист делает в среднем 5,3 клевков в мин, из них успешных — 4,0. При кормёжке головастиками и молодыми лягушками на мелководье в пойме р. Сава в Хорватии частота клевков составляла 5,9 в мин, из них 2,9 — успешных (Schulz, 1998).
Добычу птица обнаруживает чаще всего визуально. Иногда в мутной воде на мелководье белые аисты используют также тактолокацию, аналогично аистам рода Mycteria (Luhrl, 1957, Резанов, 2001). По наблюдениям А. Г. Резанова (2001) на юге Украины, зондирование мутной воды и илистого дна осуществлялось безостановочно слегка приоткрытым клювом. Аисты ходили по мелководью, делая по 43-89 шагов в мин, постоянно прощупывая участки дна перед собой. 98,9% клевков были разовыми тактильными зондированиями. Успешность кормёжки составила 2,3%.
Аисты могут поедать и мёртвых животных, например, снулую рыбу или погибших во время сенокошения птенцов, и даже питаться отбросами. В Испании в 1990-х гг. они освоили свалки и теперь кормятся там вместе с чайками и врановыми. Часть птиц даже зимует на свалках (Martin, 2002, Tortosa et al., 2002).
Описаны случаи клептопаразитизма. Так, однажды наблюдали аиста, который преследовал в воздухе серую ворону, пытаясь отобрать пойманную мышь. Предполагается, что такое поведение связано с недостатком пищи (Creutz, 1988). Могут аисты отбирать добычу и у озёрных чаек (Ranner, Szinovatz, 1987).
Кормятся аисты как поодиночке, так и стаями. В местах, богатых кормом, могут образовываться огромные скопления, которые на зимовках иногда достигают десятков тысяч особей. Причём в скоплениях эффективность кормления аистов повышается, поскольку они лучше защищены от хищников и меньше времени тратят на осматривание (Carrascal et al., 1990).
В гнездовой период аисты добывают корм, как правило, поблизости от гнёзда, но могут летать за пищей и за несколько километров. От расстояния к основным кормовым угодьям в значительной степени зависит успешность размножения. Исследования на Эльбе в Германии показали, что среднее расстояние от гнезда до мест сбора корма обратно пропорционально количеству выращенных птенцов (Dziewiaty, 1999). Обнаружена достоверная корреляция между числом вылетевших птенцов и долей влажных лугов, болот и водоёмов на гнездовой территории (Nowakowski, 2003). По данным наблюдений за одним из гнёзд в Силезии в Польше, птицы летали за едой чаще всего к нескольким предпочитаемым местам, расположенным на расстоянии от 500 до 3 375 м, в среднем 1 900 м (Jakubiec, Szymocski, 2000). Наблюдения за другой парой в Померании на севере Польши показали, что аисты кормились на территории около 250 га. Больше чем в половине случаев они искали добычу на нескольких предпочитаемых участках, которые составляли лишь 12% общей площади. 65% времени они кормились на лугах и пастбищах, 24% — на полях и 11% — на пруду. Максимальное расстояние полётов за добычей — 3 600 м, среднее — 826 м. В 53% случаев аисты кормились не далее 800 м от гнезда. Дальше всего они летали, когда уже подросли птенцы. Интересно, что самец и самка различались в своих предпочтениях, кормясь преимущественно в разных местах (Oigo, Bogucki, 1999). На Эльбе в 80% случаев аисты собирали корм не далее 1 км от гнезда (Dziewiaty, 1992). Максимальное расстояние полётов за кормом, определённое для окольцованных птиц в Зап. Европе, составляет 10 км (Lakeberg, 1995).
Анализ 242 образцов пищи, собранных в негнездовой период на Украине, показал, что весной наибольшее значение имеют земноводные и щитни, в августе — прямокрылые и различные жуки. Птенцов аисты выкармливают в основном земноводными и насекомыми на различных стадиях развития. Из насекомых наибольшее значение имеют прямокрылые и жуки, всего в питании обнаружены представители 19 семейств 3 отрядов (Смогоржевський, 1979).
В погадках, собранных в верховьях Киевского вдхр. в Черниговской обл., 96,1% фрагментов от общего числа принадлежали остаткам членистоногих. Причём питание аистов было очень разнообразным: в одной погадке находили до 130 видов животных, в том числе таких мелких, как муравьи. Среди насекомых преобладали жесткокрылые (35,3%), перепончатокрылые (21,0%) и ручейники (19,6%). Позвоночные в питании играли лишь незначительную роль (Марисова, Самофалов, Сердюк, 1992).
По данным анализа 337 погадок, собранных в южной и центральной частях Белоруссии в 1986-1992 гг., основу питания белого аиста составляли беспозвоночные — 99% общего числа определённых экземпляров кормовых объектов. Преобладали водные жуки и клопы, массовые виды жужелиц, населяющие преимущественно увлажнённые местообитания, моллюски. В населённых пунктах повышается доля мелких млекопитающих и насекомых, характерных для сухих биотопов (Самусенко, 1994). М. И. Лебедева (1960) в погадках, собранных в Беловежской пуще, обнаружила среди 187 кормовых объектов 80 экз. моллюсков, 75 — насекомых, 24 лягушки, 8 живородящих ящериц. Из насекомых найдены 42 стрекозы, 20 личинок жуков-плавунцов и водолюбов, 9 медведок, 2 кузнечика, 1 гусеница. По данным А.П. Крапивного (1957), в питании птенцов белого аиста в Беловежской пуще 72,5% по весу составляли позвоночные, из них 60,6% — лягушки. Доля дождевых червей была равной всего 1%.
В Калужской обл. энтомологический анализ погадок показал наличие представителей 17 видов, относящихся к 7 семействам отряда жесткокрылых (Coleoptera). Наиболее обычными оказались представители семейства жужелиц (Carabidae) — 41%. Далее идут пластинчатоусые жуки (Scarabaeidae) — 22%, водолюбы (Hydrophilidae) — 15%, листоеды (Chrysomelidae) и стафилины (Staphylinidae) — по 7%, плавунцы (Dytiscidae) и ложнослоники (Anthribidae) — по 4%. Представленные виды жуков в основном являлись обитателями умеренно влажных и сухих лугов, а также антропогенных ландшафтов, и были характерны для поверхности почвы — 44%, населяли небольшие пруды и лужи или являлись навозниками — по 19%, за ними следовали жесткокрылые, населяющие поля и обитающие на растительности, а также населяющие смешанные леса и живущие на коре и листьях — по 7%. В Тверской обл. в пище отмечены представители 7 семейств жуков, большинство из которых составляют пластинчатоусые и жужелицы (61,3%) (Николаев, 2000).
В Мазурии в Польше из 669 собранных погадок 97,3% содержали остатки насекомых (преобладали представители семейств Carabidae, Silphidae, Dytiscidae, Scarabeidae), 72,2% — мелких млекопитающих (в основном кроты, мыши и полёвки), 1,6% — моллюсков, 1,0% — мелких птиц, 0,7% — земноводных. Доля насекомых в питании была наибольшей на полях во время роста зерновых и люцерны и на скошенных лугах и полях после уборки урожая, довольно высокой — на вспаханных полях (Pinowski et al., 1991). В Австрии во время гнездового периода по количеству в пище преобладали прямокрылые (67,7%) и жуки (24,1%), по весу — позвоночные (55,5%), прежде всего мелкие грызуны (33,2%). Среди насекомых аисты предпочитали саранчовых, жужелиц, листоедов и пластинчатоусых жуков. В апреле-июне рацион был более разнообразным, с преобладанием мелких грызунов, в июле-августе преобладали прямокрылые (Sackl, 1987). В питании летних стай неразмножающихся птиц на лугах в Польше по количеству преобладали насекомые (83%), прежде всего жуки, по биомассе — мелкие млекопитающие, в основном полёвки (58%), насекомые (22%) и дождевые черви (11,5%) (Antczak et al., 2002). Исследования в Греции показали широкую вариабельность рациона в различных местообитаниях, но в погадках везде преобладали остатки насекомых, прежде всего прямокрылых и жуков (Tsachalidis, Goutner, 2002).
Рацион аистов может изменяться по годам, в зависимости от погодных условий. Так, на севере Германии в 1990 г., когда отмечался всплеск численности мышевидных грызунов, последние составляли на двух участках, где проводились исследования, 59,1 и 68% веса пищи, а в 1991 г. — всего лишь 3,6 и 3,8%. В очень влажный 1991 г. в питании преобладали дождевые черви — 50 и 61,6% по весу (Thomsen, Struwe, 1994). На юге Германии в разные годы весовая доля дождевых червей в питании белого аиста колебалась от 28,9 до 84%, членистоногих — от 8,9 до 28,5%, пиявок — 0 до 51,9%, грызунов — от 1,5 до 55,2%, лягушек — от 1,2 до 5,4% (Lakeberg, 1995).
Одна из основных групп насекомых, которыми питается белый аист — прямокрылые, прежде всего саранча. Наибольшее значение в рационе она имеет на местах зимовки в Африке, в связи с чем в языках некоторых африканских народов белый аист называется «саранчовой птицей». Аисты могут поедать огромное количество саранчи, объедаясь иногда так, что не могут взлететь. Во время налёта саранчи на Хортобадь в Венгрии в 1907 г. в пищеварительном тракте одного из добытых аистов обнаружили около 1 000 экз. саранчи. Желудок и пищевод птицы были набиты до самой глотки. В одной из аистиных погадок нашли 1 600 мандибул саранчи (Schenk, 1907). По оценке последнего автора, стая из 100 аистов способна за день уничтожить 100 тыс. экз. этих опасных вредителей. В местах гнездования белый аист также уничтожает большое количество вредителей сельского хозяйства, прежде всего медведок (Gryllotalpa gryllotalpa), долгоносиков, проволочников. По данным А.П. Крапивного (1957), в Беловежской пуще в питании птенцов медведки составляли 8% по количеству и почти 14% по весу. В Мазурском Поозерье в Польше 31% погадок содержал остатки проволочников, 14% — долгоносиков, 16% — медведок (Pinowska et al., 1991). В Зап. Франции в пище, которую аисты приносили птенцам, преобладали жуки-водолюбы и медведки (Barbraud, Barbraud, 1998).
При содержании в неволе суточная потребность в пище взрослого аиста колеблется от 300 г в тёплое время года до 500 г зимой. В год птице необходимо 110-130 кг (Bloesch, 1982). Суточная потребность в энергии пары аистов, выкармливающих птенцов, оценивается в 4 660 кДж. Такое её количество даёт потребление 1,4 кг дождевых червей, 1 044 г лягушек или 742 г мелких грызунов (Profus, 1986). По другим данным, пара с 1-2 птенцами потребляет около 5200 кДж (B5hning-Gaese, 1992). На р. Сава в Хорватии пара аистов ежедневно приносила птенцам в возрасте 3-6 недель в среднем 1,4 кг пищи на птенца (Schulz, 1998), на севере Германии (возраст птенцов 3-8 недель) — 1,2 кг (Struwe, Thomsen, 1991).
Для белого аиста наиболее выгодной в энергетическом плане пищей являются позвоночные животные. Во влажных местообитаниях это обычно земноводные. Однако из-за мелиоративных и гидротехнических работ численность их во многих странах значительно снизилась. Так, питание находившейся под наблюдением пары аистов в швейцарской Юре на 2/3 состояло из дождевых червей, позвоночные составляли лишь 0,4% (Wermeille, Biber, 2003). В таких условиях все большее значение для аистов приобретают грызуны. Наблюдения в долине р. Обры на западе Польши показали, что успешность размножения и даже количество заселённых гнёзд были выше в годы с высокой численностью обыкновенной полёвки (Microtus arvalis) (Tryjanowski, Kuzniak, 2002).
Враги, неблагоприятные факторы
Естественных врагов у белого аиста мало. Гнёзда могут разорять крупные хищные птицы, врановые, куницы. Взрослые птицы становятся жертвами нападений орлов, орланов, крупных четвероногих хищников — лисиц, бродячих собак, волков и т.п. Однако гибель большинства взрослых белых аистов прямо или косвенно связана с человеком.
В наибольшей степени за смертность ответственны линии электропередачи. В 1986-1989 гг. на Украине из 489 случаев гибели взрослых аистов с известной причиной 64,0% пришлось на ЛЭП. Среди жертв электролиний 80,8% погибли на опорах от поражения током и 19,2% разбились о провода. Наибольшую опасность электролинии представляют для молодых плохо летающих птиц: 72,8% случаев гибели приходится на аистов, недавно покинувших гнездо. На втором месте оказалось прямое уничтожение людьми — 12,7%. 8,8% аистов погибли в результате драк на гнёздах и при формировании стай перед отлётом, 7,6% — из-за неблагоприятных погодных условий, 2,9% — из-за отравления пестицидами, 1,6% — из-за столкновений с транспортом, 1,2% — из-за болезней, 0,8% — от хищников, 0,4% — из-за падения в большие трубы. Таким образом, всего по не связанным с деятельностью человека причинам погибли лишь 18,4% аистов. Основная причина гибели птенцов (742 случая с известной причиной) — выбрасывание птенцов родителями из гнёзд. На неё приходится 41,9%. 20,2% птенцов погибли из-за неблагоприятных погодных условий, 12,9% — из-за падения гнёзд, 7% — при драках между взрослыми аистами на гнёздах, 6,2% — уничтожены человеком, 4,5% — из-за сгорания гнёзд, 2,7% — в результате гибели родителей, 2,0% погибли от хищников, 1,5% — отравились, 1,1% — погибли из-за принесённых в гнездо материалов (Грищенко, Габер, 1990).
В Калужской обл. картина несколько иная. По данным, собранным в 1960-99 гг., основной причиной гибели взрослых птиц служит браконьерский отстрел. На его долю приходится 74% случаев с установленной причиной гибели (п = 19). В 21% случаев птицы гибли на ЛЭП, 1 раз взрослая птица погибла во время драки за гнездо с другими аистами. Основной причиной гибели слетков является контакт с электрическими коммуникациями: от удара током на открытых трансформаторах и опорах ЛЭП, а также при столкновении с проводами. Часть случаев пропажи молодых птиц вскоре после вылета из гнёзд, вероятно, следует отнести к браконьерскому отстрелу. Связаны такие различия с тем, что на территориях, недавно заселённых аистами, отношение людей к ним гораздо менее благожелательное. Известны даже случаи уничтожения появившихся гнёзд. Так, первое гнездо в Мордовии было разрушено местными жителями из-за опасений, что аисты повредят посевы огурцов (Лапшин, Лысенков, 1997). В Нижегородской обл. основная причина гибели гнёзд — преследование человеком (Бакка, Бакка, Киселева, 2000). Уничтожение взрослых птиц и разорение гнёзд отмечалось на юго-востоке Туркменистана, где аисты пытались гнездиться в 1980-х гг. (Белоусов, 1990). Однако и в тех регионах, где белый аист обитает уже давно, отношение к нему местного населения изменилось в худшую сторону. Об этом свидетельствует хотя бы высокий процент уничтожения птиц людьми среди причин гибели и разрушение гнёзд на опорах линий электропередач.
Среди причин гибели птенцов, как уже упоминалось выше, на первом месте стоит родительский инфантицид. Существенная часть птенцов выбрасывается из гнёзд или даже съедается взрослыми аистами. Так, в Беловежской пуще выбрасывали птенцов почти 30% пар, причём иногда уничтожались даже все птенцы выводка (Федюшин, Долбик, 1967). В Испании инфантицид отмечен в 18,9% находившихся под наблюдением гнёзд. Во всех случаях выбрасывался самый слабый птенец. Средний возраст выброшенных аистят—7,3 дня (Тоrtosa, Redondo, 1992). Обычно такое поведение связывают с нехваткой корма. По Д. Лэку (1957), инстинкт абортирования части отложенных яиц или вылупившихся птенцов — приспособление, позволяющее привести размер семьи в соответствие с количеством наличной пищи. Предполагается, что распространённость инфантицида у белого аиста связана с отсутствием сиблицида и конкуренции за пищу в выводках. Родители приносят большое количество мелкого корма, и более крупные птенцы не могут монополизировать его. Поскольку наиболее слабые птенцы не погибают сами, их «приходится» уничтожать родителям (Тог-tosa, Redondo, 1992, Zielicski, 2002).
Аналогичная ситуация отмечается не только на территории бывш. СССР, но и в других странах. Больше всего взрослых аистов погибает на ЛЭП, наибольшую опасность линии электропередач представляют для молодых, ещё плохо летающих птиц. Это отмечено в Болгарии (Нанкинов, 1992), Германии (Riegel, Winkel, 1971, Fiedler, Wissner, 1980), Испании (Garrido, Femandez-Cruz, 2003), Польше (Jakubiec, 1991), Словакии (Fulin, 1984), Швейцарии (Moritzi, Spaar, Biber, 2001). В округе Росток в Восточной Германии из 116 погибших птенцов белого аиста 55,2% были выброшены родителями, 20,7% погибли из-за падения гнёзд, 9,5% — от переохлаждения (Zollick, 1986). На путях пролёта и в местах зимовки основными причинами гибели аистов служат отстрел и другие формы преследования людьми, гибель на ЛЭП, отравление пестицидами (Schulz, 1988). Если многотысячный поток мигрирующих аистов пересекает район с густой сетью электролиний, одновременно гибнут десятки особей (Нанкинов, 1992).
Во многих странах Африки белый аист — традиционно охотничий вид. По данным возвратов колец, в Сев. и Зап. Африке около 80% случаев гибели приходится на отстрел. По подсчётам X. Шульца (1988), в 1980-х гг. на восточном пролётном пути ежегодно отстреливалось 5-10 тыс. аистов, из них 4-6 тыс. — в Ливане.
Массовая гибель аистов может вызываться катастрофическими погодными явлениями — бурями, крупным градом и т.п. 5 августа 1932 г. возле одного из сел на севере Болгарии во время небывалого градобоя (с неба падали куски льда до полукилограмма весом!) погибло около 200 аистов и ещё около сотни остались с поломанными ногами и крыльями (Schumann, 1932). В 1998 г. в двух сёлах Львовской обл. почти все птенцы в 19 находившихся под наблюдением гнёздах погибли во время сильных дождей (Горбулшська и др., 2004). Большой урон может причинять возвращение холодов после прилёта аистов. Так, в 1962 г. во Львовской обл. сотни особей погибли из-за морозов и снегопадов в третьей декаде марта (Черкащенко, 1963).
Иногда птенцы погибают, пытаясь проглотить принесённую родителями слишком крупную добычу. Например, отмечен случай гибели аистёнка, подавившегося ужом (Kuppler, 2001). Опасность птенцам представляют и некоторые принесённые родителями в гнездо материалы — куски шпагата, пакли, в которых аистята могут запутываться, обрывки плёнки или клеёнки в лотке, на которых собирается вода.
Белый аист может становиться жертвой клептопаразитизма. Например, на Днепре в Черкасской обл. наблюдали случай нападения на пролетавшего над рекой аиста орлана-белохвоста (Haliaeetus albicilla). Аист отрыгнул добычу, две рыбёшки из которой орлан подобрал с поверхности воды (Лопарев, 1997).
К неблагоприятным факторам принадлежат происходящие в последние десятилетия изменения среды обитания. Постройки с соломенными и тростниковыми крышами, на которых охотно гнездились аисты, практически исчезли из деревень. Уменьшается и количество пригодных для гнездования старых деревьев в населённых пунктах. Чрезмерная мелиорация, затопление пойм рек водохранилищами, нарушение нормального гидрорежима водоёмов приводит к обеднению кормовой базы. Особенно актуально это для многих стран Зап. Европы, где приходится специально разводить земноводных на корм аистам. В последнее время добавилась ещё одна проблема — сокращение площадей традиционно использующихся лугов и пастбищ во многих регионах Вост. Европы и Сев. Азии из-за экономического спада. Всё увеличивающаяся химизация сельского хозяйства вызывает накопление пестицидов в цепях питания, что вызывает отравления и заболевания птиц. В наибольшей степени это проявляется на местах зимовки, где проводится активная борьба с саранчой и другими вредителями сельского хозяйства, которые служат основным кормом аистов.
В Средней Азии наиболее важным фактором, повлиявшим на изменения ареала и численности, стало освоение новых земель под посевы сельскохозяйственных культур с преобладанием монокультуры хлопчатника, вырубка деревьев в долинах рек, осушение болот, уменьшение площади рисовых полей. Благодаря укрупнению полей было вырублено много лесополос. Современная архитектура и градостроительные тенденции не способствуют гнездованию белого аиста в населённых пунктах (Сагитов, 1990, Шерназаров и др., 1992).
В России существенным фактором, ограничивающим численность гнездящихся пар, служит разрушение гнёзд на церквях в связи с их реставрацией, на телеграфных столбах и опорах ЛЭП при обслуживании электрических коммуникаций, а также демонтаж водонапорных башен для установки на новое место или сдачи на металлолом. Последний фактор представляется особенно угрожающим, так как на водонапорных башнях гнездится более половины российской группировки белого аиста.
К неблагоприятным факторам следует отнести и ухудшение положительного отношения к белому аисту местного населения, утрата давних народных традиций. Так, проведённый в Киевской обл. опрос показал, что значительная часть сельских жителей не только не знает, как привлечь белого аиста на гнездование, но и не хочет иметь гнездо на усадьбе (Грищенко и др., 1992). Это при том, что наличие гнезда раньше считалось большим благом, привлечение белого аиста на гнездование было одним из элементов древней аграрной магии (Грищенко, 19986, 2005). В Узбекистане белый аист считался священной птицей, но теперь население в некоторых местах занимается разорением гнёзд и сбором яиц (Сагитов, 1990).
На юге Украины у белого аиста зарегистрированы 4 вида гельминтов: Dyctimetra discoidea, Chaunocephalus ferox, Tylodelphys excavata, Histriorchis tricolor (Корнюшин и др., 2004).
В гнёздах белого аиста обнаружено около 70 представителей разных видов насекомых, главным образом жуков (Coleoptera) (Hicks, 1959).
Хозяйственное значение, охрана
Белый аист уничтожает большое количество вредителей сельского хозяйства, прежде всего насекомых и грызунов. Он широко известен как один из наиболее активных истребителей саранчи. Аист может приносить некоторый ущерб рыбному и охотничьему хозяйству, поедая рыбу, птенцов, зайчат и т.п., однако подобное носит лишь случайный характер, и такие кормовые объекты не занимают сколько-нибудь заметного места в рационе белого аиста. Более или менее существенный ущерб рыбному хозяйству возникает лишь там, где образуются большие скопления аистов и практически отсутствует другая пища (например, на рыборазводных хозяйствах в Израиле). В странах же Вост. Европы и Сев. Азии это случается редко.
Белый аист — давний спутник человека, он имеет огромное эстетическое значение, считается одной из наиболее любимых и почитаемых у многих народов птиц. Культ его сформировался в глубокой древности, вполне вероятно вскоре после возникновения производящего хозяйства (Грищенко, 19986, 2005). Аист — прекрасный объект для экологического образования и воспитания, принимает помощь человека, позитивно воздействует на эмоции живущих рядом людей. Для охраны аиста необходима активная пропагандистская и разъяснительная работа, возрождение давних народных традиций помощи этой птице. При этом благодаря большой популярности белого аиста удаётся привлечь к природоохранной деятельности значительное количество людей. Большую эффективность имеют широкомасштабные научно-пропагандистские акции, например, проводившиеся на Украине операции «Лелека» («Аист») и «Год белого аиста» (Грищенко, 1991,19966, Грищенко и др., 1992). Особенно важны как пропагандистская работа, так и практическая помощь в зоне расселения — для закрепления птиц на новых местах гнездования.
Белый аист занесён в Красные книги Казахстана, Узбекистана, а в Российской Федерации — в Красные книги Карелии, Мордовии, Чечни, Краснодарского и Ставропольского краёв, Белгородской, Волгоградской, Калужской, Кировской, Липецкой, Московской, Нижегородской, Пензенской, Ростовской, Рязанской, Тамбовской, Тверской и некоторых других областей.
Физические характеристики
Плотно сбитое тело аиста белого 100-115 см от кончика клюва до конца хвоста, вес 2,5 — 4,4 кг, размах крыльев 195 — 215 см. У крупной болотной птицы белое оперение тела, черные полётные перья на крыльях. Пигмент меланин и каротиноиды в рационе питания аистов обеспечивают черный окрас.
p, blockquote 4,0,0,0,0 -->
У взрослых белых аистов длинные заостренные красные клювы, длинные красные лапы с частично перепончатыми пальцами и длинная тонкая шея. У них черная кожа вокруг глаз, когти тупые и похожи на гвозди. Самцы и самочки выглядят одинаково, самцы немного больше. Перья на груди длинные и образуют своеобразную накладку, которую птицы используют при ухаживании.
p, blockquote 5,0,0,0,0 -->
На длинных и широких крыльях белый аист легко парит в воздухе. Птицы взмахивают крыльями медленно. Как и большинство водоплавающих пернатых, парящих в небе, белые аисты выглядят зрелищно: длинные шеи вытянуты вперед, а длинные лапы вытянуты назад далеко за край короткого хвоста. Они машут огромными, широкими крыльями не часто, сохраняют энергию.
p, blockquote 6,0,0,0,0 -->
На земле белый аист идет медленным ровным шагом, вытянув голову вверх. В состоянии покоя склоняет голову к плечам. Первичные лётные перья ежегодно линяют, в период размножения вырастает новое оперение.
p, blockquote 7,1,0,0,0 -->
Какие места предпочитают белые аисты для жилья
Местами обитания белый аист выбирает:
p, blockquote 8,0,0,0,0 -->
- берега рек,
- болота,
- каналы,
- луга.
Белые аисты избегают областей, заросших высокими деревьями и кустарниками.
p, blockquote 9,0,0,0,0 -->
Белый аист в полете
Диета аиста
Белый аист активен в течение дня, предпочитает кормиться в мелких водно-болотных и сельскохозяйственных угодьях, на травяных лугах. Белый аист является хищником и питается:
p, blockquote 10,0,0,1,0 -->
- амфибиями,
- ящерицами,
- змеями,
- лягушками,
- насекомыми,
- рыбами,
- мелкими птицами,
- млекопитающими.

p, blockquote 11,0,0,0,0 -->